2. Энергетический обмен (катаболизм, диссимиляция)
Универсальным источником энергии во всех клетках служит АТФ (аденозинтрифосфат, или аденозинтрифосфорная кислота).
Все энергетические затраты любой клетки обеспечиваются за счёт универсального энергетического вещества — АТФ.
АТФ синтезируется в результате реакции фосфорилирования, то есть присоединения одного остатка фосфорной кислоты к молекуле АДФ (аденозиндифосфата):
АДФ + h4PO4+ 40 кДж = АТФ + h3O.
Энергия запасается в форме энергии химических связей АТФ. Химические связи АТФ, при разрыве которых выделяется много энергии, называются макроэргическими.
При распаде АТФ до АДФ клетка за счёт разрыва макроэргической связи получит приблизительно \(40\) кДж энергии.
Энергия для синтеза АТФ из АДФ выделяется в процессе диссимиляции.
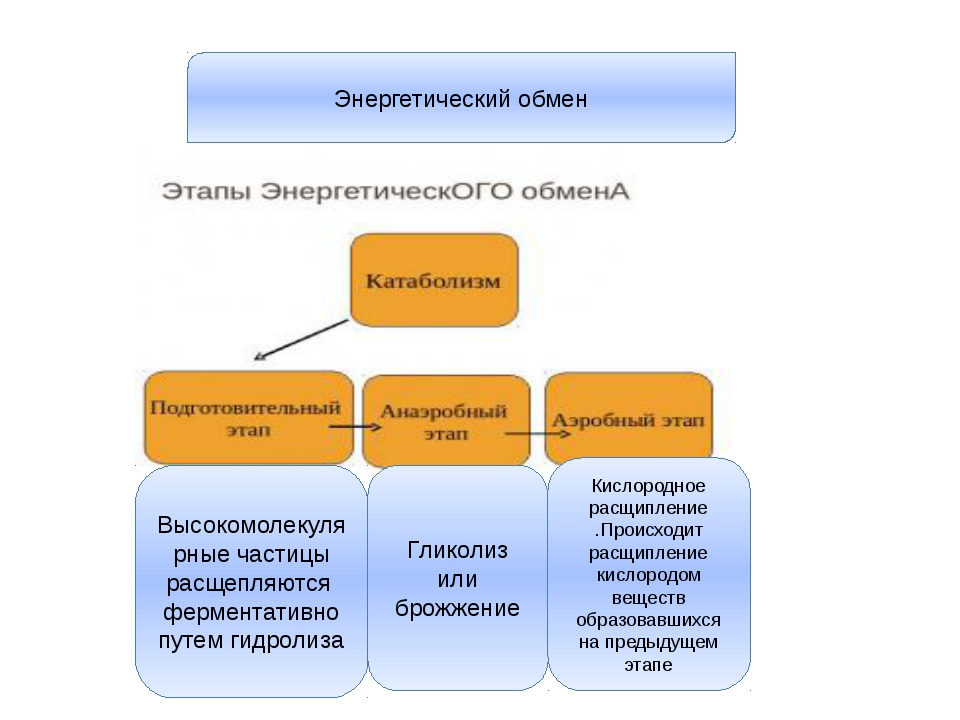
Энергетический обмен (диссимиляция, катаболизм) — это совокупность химических реакций постепенного распада органических соединений, сопровождающихся высвобождением энергии, часть которой расходуется на синтез АТФ.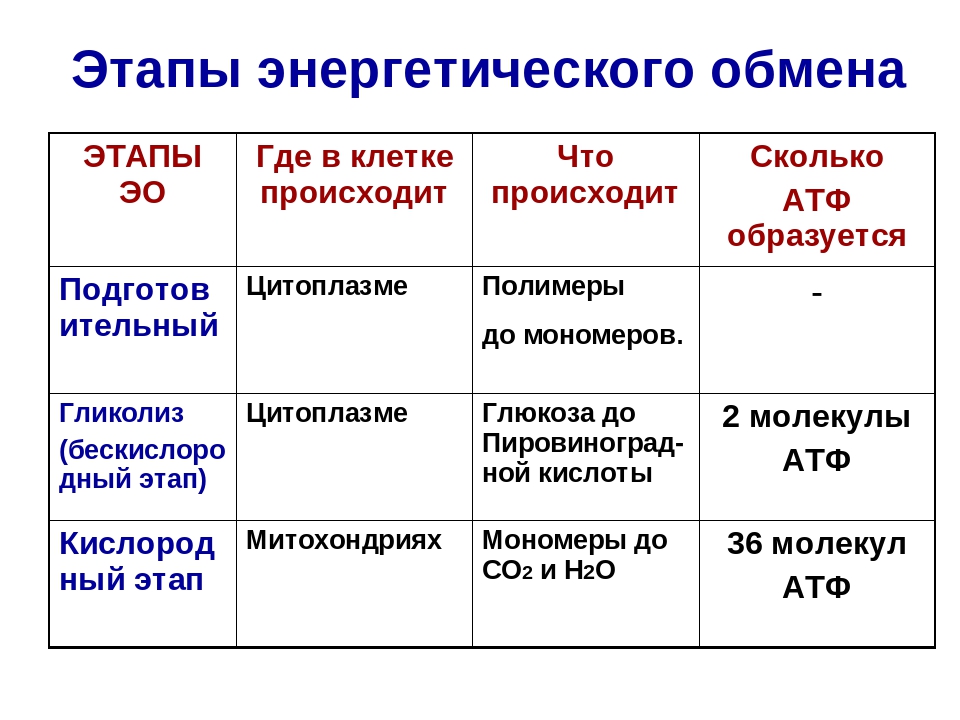
В зависимости от среды обитания организма, диссимиляция может проходить в два или в три этапа.
Процессы расщепления органических соединений у аэробных организмов происходят в три этапа: подготовительный, бескислородный и кислородный.
В результате этого органические вещества распадаются до простейших неорганических соединений.
У анаэробных организмов, обитающих в бескислородной среде и не нуждающихся в кислороде (а также у аэробных организмов при недостатке кислорода), диссимиляция происходит в два этапа: подготовительный и бескислородный.
В двухэтапном энергетическом обмене энергии запасается гораздо меньше, чем в трёхэтапном.
Первый этап — подготовительный
Подготовительный этап заключается в распаде крупных органических молекул до более простых: полисахаридов — до моносахаридов, липидов — до глицерина и жирных кислот, белков — до аминокислот.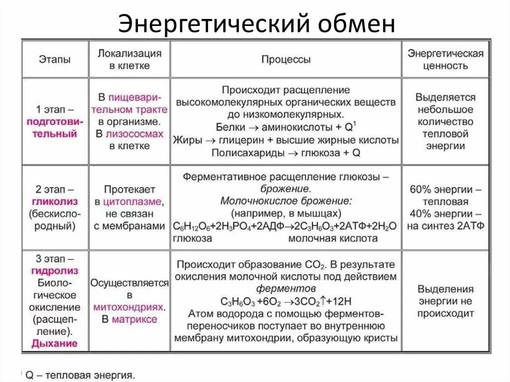
Этот процесс называется пищеварением. У многоклеточных организмов он осуществляется в желудочно-кишечном тракте с помощью пищеварительных ферментов. У одноклеточных организмов — происходит под действием ферментов лизосом.
В ходе биохимических реакций, происходящих на этом этапе, энергии выделяется мало, она рассеивается в виде тепла, и АТФ не образуется.
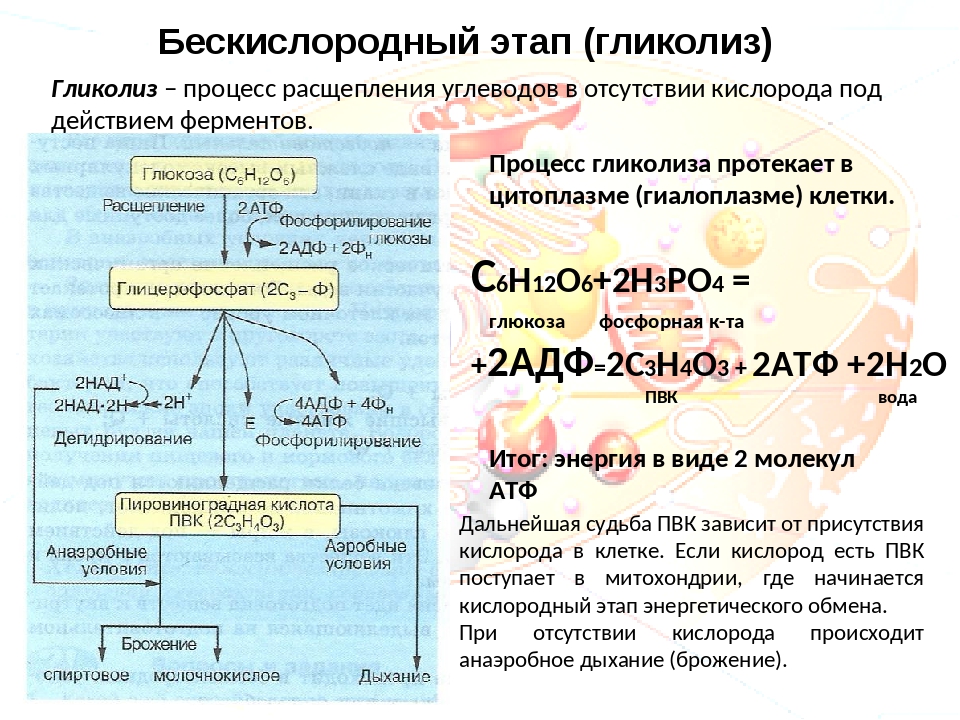
Второй этап — бескислородный (гликолиз)
Второй (бескислородный) этап заключается в ферментативном расщеплении органических веществ, которые были получены в ходе подготовительного этапа. Кислород в реакциях этого этапа не участвует.
Биологический смысл второго этапа заключается в начале постепенного расщепления и окисления глюкозы с накоплением энергии в виде \(2\) молекул АТФ.
Процесс бескислородного расщепления глюкозы называется гликолиз.
Гликолиз происходит в цитоплазме клеток.
Он состоит из нескольких последовательных реакций превращения молекулы глюкозы C6h22O6 в две молекулы пировиноградной кислоты — ПВК C3h5O3 и две молекулы АТФ (в виде которой запасается примерно \(40\) % энергии, выделившейся при гликолизе). Остальная энергия (около \(60\) %) рассеивается в виде тепла.
C6h22O6+2h4PO4+2АДФ=2C3h5O3+2АТФ +2h3O.
Получившаяся пировиноградная кислота при недостатке кислорода в клетках животных, а также клетках многих грибов и микроорганизмов, превращается в молочную кислоту C3H6O3.
HOOC−CO−Ch4пировиноградная кислота→НАД⋅H+H+лактатдегидрогеназаHOOC−CHOH−Ch4молочная кислота.
В мышцах человека при больших нагрузках и нехватке кислорода образуется молочная кислота и появляется боль. У нетренированных людей это происходит быстрее, чем у людей тренированных.
При недостатке кислорода в клетках растений, а также в клетках некоторых грибов (например, дрожжей), вместо гликолиза происходит спиртовое брожение: пировиноградная кислота распадается на этиловый спирт C2H5OH и углекислый газ CO2:
C6h22O6+2h4PO4+2АДФ=2C2H5OH+2CO2+2АТФ+2h3O.
Третий этап — кислородный
В результате гликолиза глюкоза распадается не до конечных продуктов (CO2 и h3O), а до богатых энергией соединений (молочная кислота, этиловый спирт) которые, окисляясь дальше, могут дать её в больших количествах. Поэтому у аэробных организмов после гликолиза (или спиртового брожения) следует третий, завершающий этап энергетического обмена — полное кислородное расщепление, или клеточное дыхание.
Этот этап происходит на кристах митохондрий.
Третий этап, так же как и гликолиз, является многостадийным и состоит из двух последовательных процессов — цикла Кребса и окислительного фосфорилирования.
Третий (кислородный) этап заключается в том, что при кислородном дыхании ПВК окисляется до окончательных продуктов — углекислого газа и воды, а энергия, выделяющаяся при окислении, запасается в виде \(36\) молекул АТФ (\(2\) молекулы в цикле Кребса и \(34\) молекулы в ходе окислительного фосфорилирования).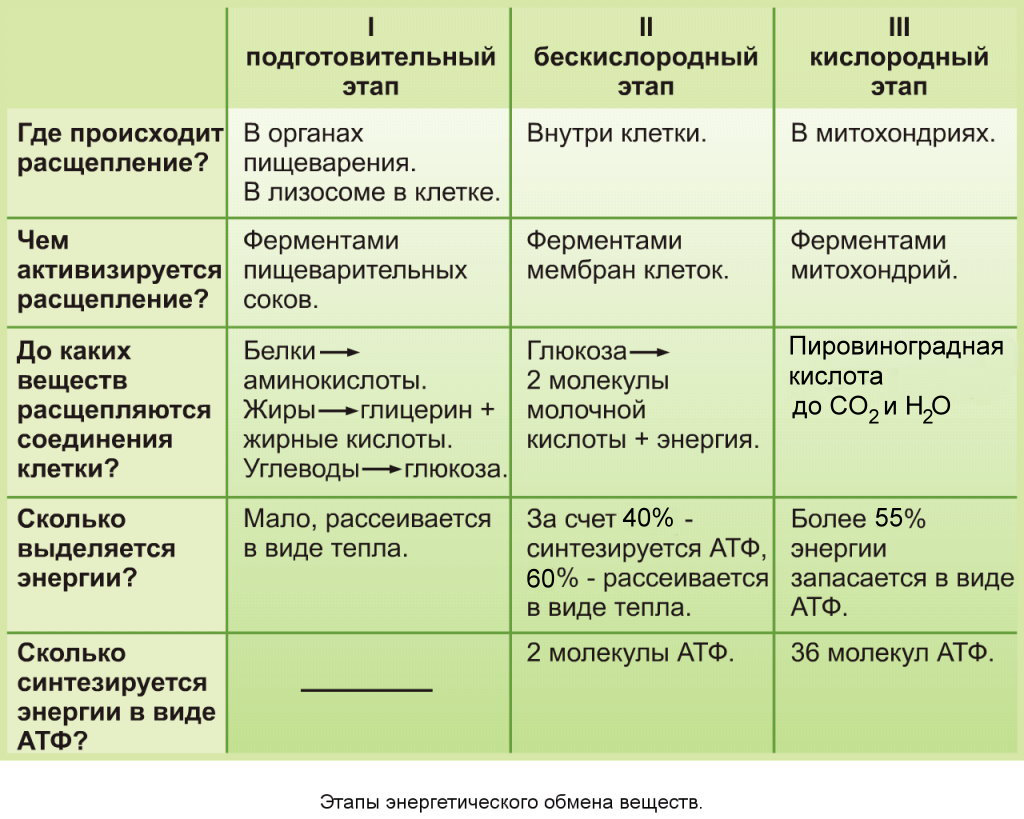
Этот этап можно представить себе в следующем виде:
2C3h5O3+6O2+36h4PO4+36АДФ=6CO2+42h3O+36АТФ.
Вспомним, что ещё две молекулы АТФ запасаются в ходе бескислородного расщепления каждой молекулы глюкозы (на втором, бескислородном, этапе). Таким образом, в результате полного расщепления одной молекулы глюкозы образуется \(38\) молекул АТФ.
Суммарная реакция энергетического обмена:
C6h22O6+6O2=6CO2+6h3O+38АТФ.
Для получения энергии в клетках, кроме глюкозы, могут быть использованы и другие вещества: липиды, белки. Однако ведущая роль в энергетическом обмене у большинства организмов принадлежит сахарам.
Источники:
Каменский А. А., Криксунов Е. А., Пасечник В. В. Биология. 9 класс // ДРОФА.
Каменский А. А., Криксунов Е. А., Пасечник В. В. Биология. Общая биология (базовый уровень) 10–11 класс // ДРОФА.
Лернер Г. И. Биология: Полный справочник для подготовки к ЕГЭ: АСТ, Астрель.
Этапы энергетического обмена
☰
Энергетический обмен — это по-этапный распад сложных органических соединений, протекающий с выделением энергии, которая запасается в макроэргических связях молекул АТФ и используется потом в процессе жизнедеятельности клетки, в том числе на биосинтез, т.е. пластический обмен.
В аэробных организмах выделяют три последовательных этапа энергетического обмена:
- Подготовительный — расщепление биополимеров до мономеров.
- Бескислородный — гликолиз — расщепление глюкозы до пировиноградной кислоты.
- Кислородный — расщепление пировиноградной кислоты до углекислого газа и воды.
Подготовительный этап
На подготовительном этапе энергетического обмена происходит расщепление поступивших с пищей органических соединений на более простые, обычно мономеры.
Хотя при этом выделяется энергия, она не запасается в АТФ и, следовательно, не может быть использована впоследствии. Энергия рассеивается в виде тепла.
Расщепление полимеров у многоклеточных сложноорганизованных животных протекает в пищеварительном тракте под действием выделяющихся сюда железами ферментов. Затем образовавшиеся мономеры всасываются в кровь в основном через кишечник. Уже кровью питательные вещества разносятся по клеткам.
При этом не все вещества разлагаются до мономеров в пищеварительной системе. Расщепление многих происходит непосредственно в клетках, в их лизосомах. У одноклеточных организмов поглощенные вещества попадают в пищеварительные вакуоли, где и перевариваются.
Образовавшиеся мономеры могут использоваться как для энергетического, так и пластического обмена. В первом случае они расщепляются, во-втором – из них синтезируются компоненты самих клеток.
Бескислородный этап энергетического обмена
Бескислородный этап протекает в цитоплазме клеток и в случае аэробных организмов включает только гликолиз — ферментативное многоступенчатое окисление глюкозы и ее расщепление до пировиноградной кислоты, которую также называют пируватом.
Молекула глюкозы включает шесть атомов углерода. При гликолизе она расщепляется до двух молекул пирувата, который включает три атома углерода. При этом отщепляется часть атомов водорода, которые передаются на кофермент НАД, который, в свою очередь, потом будет участвовать в кислородном этапе.
Часть выделяющейся при гликолизе энергии запасается в молекулах АТФ. На одну молекулу глюкозы синтезируется всего две молекулы АТФ.
Энергия, оставшаяся в пирувате, запасенная в НАД, у аэробов далее будет извлечена на следующем этапе энергетического обмена.
В анаэробных условиях, когда кислородный этап клеточного дыхания отсутствует, пируват «обезвреживается» в молочную кислоту или подвергается брожению. При этом энергия не запасается. Таким образом, здесь полезный энергетический выход обеспечивается только малоэффектвным гликолизом.
При этом энергия не запасается. Таким образом, здесь полезный энергетический выход обеспечивается только малоэффектвным гликолизом.
Кислородный этап
Кислородный этап протекает в митохондриях. В нем выделяют два подэтапа: цикл Кребса и окислительное фосфорилирование. Поступающий в клетки кислород используется только на втором. В цикле Кребса происходит образование и выделение углекислого газа.
Цикл Кребса протекает в матриксе митохондрий, осуществляется множеством ферментов. В него поступает не сама молекула пировиноградной кислоты (или жирной кислоты, аминокислоты), а отделившаяся от нее с помощью кофермента-А ацетильная группа, включающая два атома углерода бывшего пирувата. За многоступенчатый цикл Кребса происходит расщепление ацетильной группы до двух молекул CO 2 и атомов водорода. Водород соединяется с НАД и ФАД. Также происходит синтез молекулы ГДФ, приводящей к синтезу потом АТФ.
На одну молекулу глюкозы, из которой образуется два пирувата, приходится два цикла Кребса. Таким образом, образуется две молекулы АТФ. Если бы энергетический обмен заканчивался здесь, то суммарно расщепление молекулы глюкозы давало бы 4 молекулы АТФ (две от гликолиза).
Таким образом, образуется две молекулы АТФ. Если бы энергетический обмен заканчивался здесь, то суммарно расщепление молекулы глюкозы давало бы 4 молекулы АТФ (две от гликолиза).
Окислительное фосфорилирование протекает на кристах – выростах внутренней мембраны митохондрий. Его обеспечивает конвейер ферментов и коферментов, образующий так называемую дыхательную цепь, заканчивающуюся ферментом АТФ-синтетазой.
По дыхательной цепи происходит передача водорода и электронов, поступивших в нее от коферментов НАД и ФАД. Передача осуществляется таким образом, что протоны водорода накапливаются с внешней стороны внутренней мембраны митохондрий, а последние ферменты в цепи передают только электроны.
В конечном итоге электроны передаются молекулам кислорода, находящимся с внутренней стороны мембраны, в результате чего они заряжаются отрицательно. Возникает критический уровень градиента электрического потенциала, приводящий к перемещению протонов через каналы АТФ-синтетазы. Энергия движения протонов водорода используется для синтеза молекул АТФ, а сами протоны соединяются с анионами кислорода с образованием молекул воды.
Энергия движения протонов водорода используется для синтеза молекул АТФ, а сами протоны соединяются с анионами кислорода с образованием молекул воды.
Энергетический выход функционирования дыхательной цепи, выраженный в молекулах АТФ, велик и суммарно составляет от 32 до 34 молекул АТФ на одну исходную молекулу глюкозы.
Этапы энергетического обмена – Российский учебник
Живой организм построен из клеток. А некоторые, например, амеба или инфузория, состоят из одной клетки. Сложные многоклеточные особи представлены сочетаниями клеток, которые образуют различные системы организма: дыхательную или пищеварительную. Весь организм пронизан клетками нервной ткани, которые обеспечивают регуляцию и управление макроорганизмом.
Живая клетка от неживой отличается тем, что в ней постоянно и непрерывно протекает два разнонаправленных процесса:
- синтез, или строительство новых органелл (пластический обмен или ассимиляция)
- катаболизм, или разрушение питательных элементов с образованием энергии (энергетический обмен или диссимиляция)
В живых особях баланс между ассимиляцией и диссимиляцией поддерживается не всегда.
Чтобы машина ехала, ей нужен бензин. А бензин делают из нефти. Чтобы макроорганизм существовал, ему нужна энергия. В учебниках биологии часто можно встретить фразу глюкоза — ресурс энергии для клетки. Это так. Но глюкоза — как нефть для автомобиля. Поэтому сначала глюкозу нужно превратить в бензин. И таким бензином для клетки будет молекула АТФ.
Молекула АТФ известна миру довольно давно. Через десять лет, в 2029 году отпразднует сто лет со дня открытия вещество, благодаря которому в живой клетке поддерживается упорядоченность структур и она может противостоять хаосу и растущей энтропии (почему так происходит, подробно рассказано на странице 30 учебника «Естествознание 11 класс» под редакцией Титова С.
В 1929 году, практически одновременно, сотрудники Института биологии кайзера Вильгельма в Берлине Карл Ломан и Отто Мейергоф и ученые Гарвардской Медицинской школы Сайрус Фиске и Йеллапрагад Суббарао в Гарвардской Медицинской школе опубликовали работы, в которых описали молекулу АТФ.
В 30 годах ХХ века в лаборатории Мейергофа появился молодой ученый, Фриц Липман, который заинтересовался энергетическими аспектами клеточного метаболизма, и в 1941 году талантливый биохимик доказал, что АТФ – основной двигатель энергетических реакций в живой клетке. А в 1953 году его вклад в физиологию совместно с Х. Кребсом был удостоен Нобелевской премии.
АТФ — аденозинтрифосфат, нуклеотид, глобальный ресурс энергии для сложных реакций обмена веществ, происходящих в клетках организма. Схематично молекула АТФ представлена на странице 99 учебника «Естествознание 11 класс» под редакцией Титова С.А..
Вся суть энергетического обмена сводится к решению задачи, как энергию из сложных молекул пищи превратить в молекулу АТФ. В процессе эволюции эта задача была решена.
В процессе эволюции эта задача была решена.
Так как же булочка из Макдональдса превращается в энергию макроэргических связей АТФ?
В энергетическом обмене выделяют несколько процессов, разделенных не только временем, но и протекающих в различных частях клетки:
- Подготовительный этап
- Гликолиз
- Кислородное окисление
Естествознание. 10 класс. Учебник
Учебник отличаются качественным современным оформлением, в нём приводятся многочисленные слайды и микрофотографии. Выполняя проблемные, поисковые и исследовательские задания, школьники не только активно усваивают материал, но и учатся мыслить, искать и анализировать информацию из разных источников, в том числе из интернета. Особое внимание уделяется практическим заданиям: ученикам предлагается проводить опыты, конструировать модели, разрабатывать проекты.
Купить
Подготовительный этап у человека и других многоклеточных макроорганизмов начинается в ротовой полости, продолжается в желудочно-кишечном тракте и представляет собой многоступенчатый процесс распада сложных полимеров белков, жиров и углеводов пищи до мономеров.
Из курса химии помним, что во время разрыва связей элементов выделяется энергия. Для образования аденозинтрифосфата этой энергии недостаточно, и она вся уходит во внешнюю среду.
Что же происходит у простых одноклеточных организмов, у которых нет ротиков и животиков? Пища, захваченная одноклеточным организмом, попадает в пищеварительную вакуоль или лизосому, где при помощи ферментов-катализаторов, способствующих пищеварению, протекает начальный этап диссимиляции.
Подготовленные во время предварительного этапа вещества переходят во второй бескислородный этап энергетического обмена, который называется гликолиз.
Два греческих слова (гликос – «сладкий» и лизис – «расщеплять») дали название второй бескислородной фазе энергетического обмена — глико́лизу.
Гликолиз является цепочкой из 10 биохимических превращений, итогом которых является трансформация стабильной молекулы глюкозы в трехуглеродные молекулы пирувата, – или пировиноградной кислоты.
Что ещё почитать?
«Подождите, – могут сказать дотошные ученики, – глюкоза у нас находится в пищеварительной системе. А процессы метаболизма идут в клетках по всему организму, например, на кончике носа или задней лапе. Как же попадает глюкоза в цитоплазму клетки во всем организме?».Глюкоза всасывается в желудочно-кишечном тракте, попадает в кровоток, током крови разносится ко всем тканям и клеткам организма, и благодаря белку-переносчику инсулину попадает в клетку.
Цитоплазма отдельной клетки – место осуществления реакций гликолиза. Энергии при этом выделяется совсем немного. Ее хватает лишь на формирование 2-х молекул АТФ. Казалось бы, энергия получена, и процесс может остановиться. Так и происходит у некоторых бактерий. Но никакому нормальному многоклеточному организму таких запасов АТФ не хватит. В пировиноградной кислоте остался еще достаточный запас энергии, которую тоже хотелось бы использовать макроорганизму.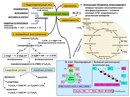
Естествознание. 11 класс. Учебник
Учебник отличаются качественным современным оформлением, в нём приводятся многочисленные слайды и микрофотографии. Выполняя проблемные, поисковые и исследовательские задания, школьники не только активно усваивают материал, но и учатся мыслить, искать и анализировать информацию из разных источников, в том числе из интернета. Особое внимание уделяется практическим заданиям: ученикам предлагается проводить опыты, конструировать модели, разрабатывать проекты.
КупитьУ многоклеточных тел пируват переходит в третью фазу диссимиляции — клеточное дыхание в митохондриях. Дыханием процесс называется, поскольку в ходе химических реакций в митохондриях происходит потребление кислорода и выделение углекислого газа в цитоплазму клетки, а дальше, с помощью кровообращения и дыхания, – во внешнюю среду.
Клеточное дыхание представлено двумя этапами:
- цикл Кребса, протекающий в матриксе митохондрий
- окислительное фосфорилирование, протекающее на кристах митохондрий при участии ферментов дыхательной цепи
Итогом кислородного этапа энергетического обмена является выделение количества энергии, достаточного для образования 36 молекул АТФ, воды и СО2. При этом нужно помнить, что аденозинтрифосфат содержит три остатка фосфата, а макроэргических связей образуется только две. Суммарное уравнение биохимических реакций, протекающих в третьей фазе диссимиляции, можно записать так:
При этом нужно помнить, что аденозинтрифосфат содержит три остатка фосфата, а макроэргических связей образуется только две. Суммарное уравнение биохимических реакций, протекающих в третьей фазе диссимиляции, можно записать так:
2C3H4O3+6O2+36H3PO4+36АДФ=6CO2+42H2O+36АТФ
В итоге этих реакций происходит накопление огромного количества энергии — 36 молекул аденозинтрифосфата против 2-х, что запасаются в процессе гликолиза. Однако поскольку эта фаза требует кислорода для своих реакций, в бескислородной среде процесс протекать не может.
При дефиците кислорода пируват окисляется до лактата. Именно ему принадлежит ощущение приятной боли после хорошей тренировки. У хорошо тренированных людей с активным кровоснабжением и хорошо развитой сетью капилляров нужно затратить большую физическую нагрузку перед тем, как начнет накапливаться молочная кислота.
Вспомним, что еще 2 молекулы аденозинтрифосфата накапливаются на этапе гликолиза. Таким образом, при распаде одной молекулы глюкозы образуется 38 молекул АТФ.
На портале LECTA В параграфе 22 учебника «Естествознание 11 класс» под редакцией Титова С.А. внимательные ученики найдут ответ на вопрос, почему цианистый калий – любимое средство убийц в детективных романах.
ТестЭнергетический обмен, подготовка к ЕГЭ по биологии
Обмен веществ
Обмен веществ (метаболизм) складывается из процессов расщепления и синтеза — диссимиляции и ассимиляции, постоянно протекающих в организме. Чтобы жизнь продолжалась, количество поступающей энергии должно превышать (или как минимум равняться) количеству расходуемой энергии, поэтому диссимиляция и ассимиляция поддерживают определенный баланс друг с другом.
Энергетический обмен
Энергетический обмен (диссимиляция — от лат.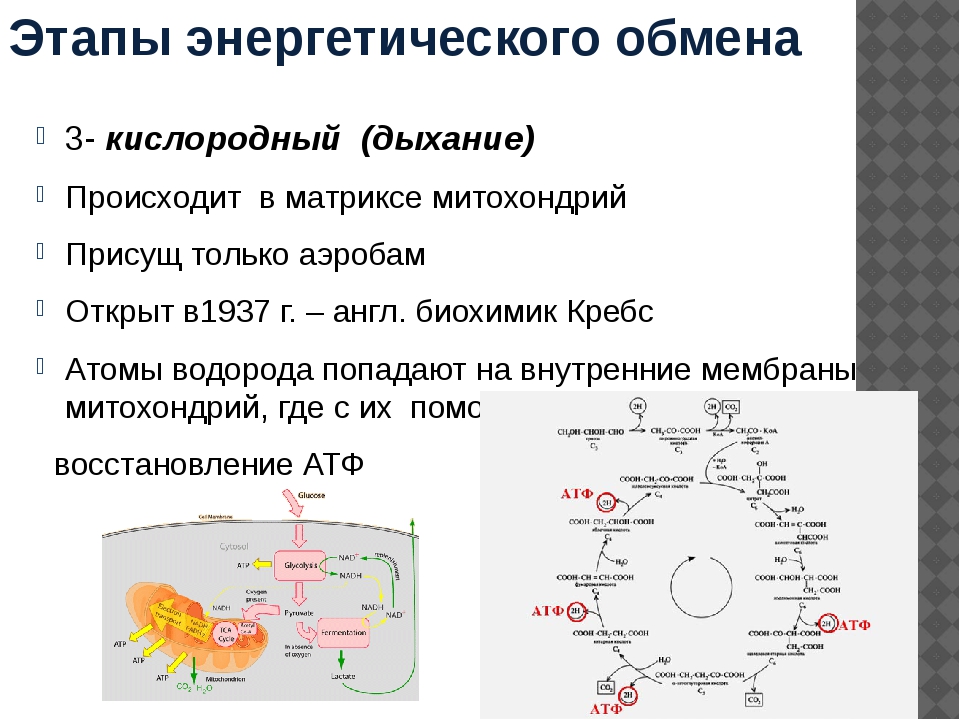 dissimilis ‒ несходный) — обратная ассимиляции сторона обмена веществ, совокупность реакций, которые приводят к высвобождению энергии химических связей. Это реакции расщепления жиров,
белков, углеводов, нуклеиновых кислот до простых веществ.
dissimilis ‒ несходный) — обратная ассимиляции сторона обмена веществ, совокупность реакций, которые приводят к высвобождению энергии химических связей. Это реакции расщепления жиров,
белков, углеводов, нуклеиновых кислот до простых веществ.
Возможно три этапа диссимиляции: подготовительный, анаэробный и аэробный. Среда обитания определяет количество этапов диссимиляции. Их может быть три, если организм обитает в кислородной среде, и два, если речь идет об организме, обитающем в бескислородной среде (к примеру, в кишечнике).
Обсудим этапы энергетического обмена более подробно:
- Подготовительный этап
- Бескислородный этап (анаэробный) — гликолиз
- Кислородный этап (аэробный)
Осуществляется в ферментами, в результате действия которых, сложные вещества превращаются в более простые: полимеры распадаются на мономеры. Это сопровождается разрывом химических связей и выделением энергии, большая часть которой рассеивается в виде тепла.
Под действием ферментов белки расщепляются на аминокислоты, жиры — на глицерин и жирные кислоты, сложные углеводы — до простых сахаров.
Этот этап является последним для организмов-анаэробов, обитающих в условиях, где кислород отсутствует. На этапе гликолиза происходит расщепление молекулы глюкозы: образуется 2 молекулы АТФ и 2 молекулы пировиноградной кислоты (ПВК). Происходит данный этап в цитоплазме клеток.
Этот этап доступен только для аэробов — организмов, живущих в кислородной среде. Из каждой молекулы ПВК, образовавшейся на этапе гликолиза, синтезируется 18 молекул АТФ — в сумме с двух ПВК выход составляет 36 молекул АТФ.
Таким образом, суммарно с одной молекулы глюкозы можно получить 38 АТФ (гликолиз + кислородный этап).
Кислородный этап протекает на кристах митохондрий (складках, выпячиваниях внутренней мембраны), где наибольшая концентрация окислительных ферментов.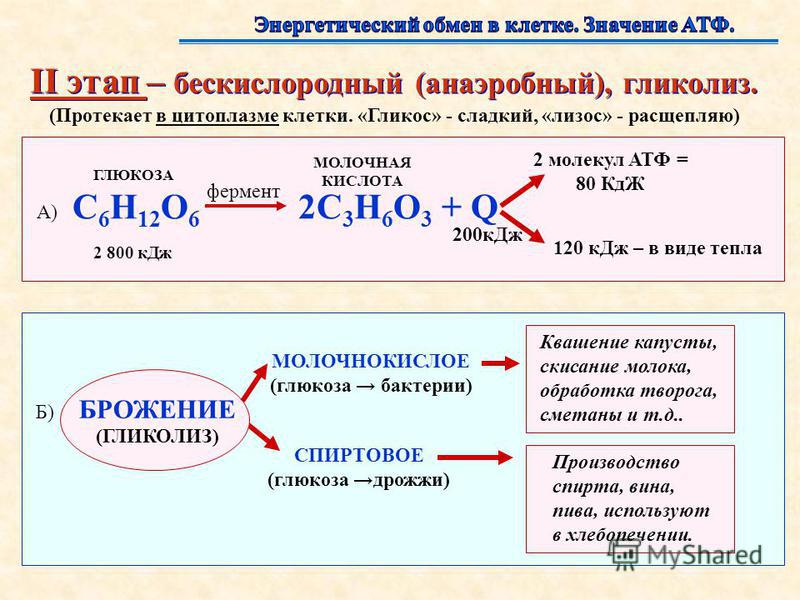 Главную роль в этом процессе играет так называемый цикл Кребса, который подробно изучает биохимия.
Главную роль в этом процессе играет так называемый цикл Кребса, который подробно изучает биохимия.
АТФ — аденозинтрифосфорная кислота
Трудно переоценить роль в клетке АТФ — универсального источника энергии. Молекула АТФ состоит из азотистого основания — аденина, углевода — рибозы и трех остатков фосфорной кислоты.
Между остатками фосфорной кислоты находятся макроэргические связи — ковалентные связи, которые гидролизуются с выделением большого количества энергии. Их принято обозначать типографическим знаком тильда «∽».
АТФ гидролизуется до АДФ (аденозиндифосфорная кислота), а затем и до АМФ (аденозинмонофосфорная кислота). Гидролиз АТФ сопровождается выделением энергии (E) на каждом этапе и может быть представлен такой схемой:
- АТФ + H2O = АДФ + H3PO4 + E
- АДФ + H2O = АМФ + H3PO4 + E
- АМФ + H2O = аденин + рибоза + H3PO4 + E
Пластический обмен
АТФ является универсальным источником энергии в клетке: энергия макроэргических связей АТФ используется для реакций
пластического обмена (ассимиляции), протекающих с затратой энергии: синтеза белка на рибосоме (трансляции),
удвоению ДНК (репликации) и т. д.
д.
В результате пластического обмена в нашем организме происходит синтез белков, жиров и углеводов.
© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Урок 24. энергетика живой клетки — Естествознание — 10 класс
Конспект на интерактивный видео-урок
по предмету «Естествознание» для «10» класса
Урок № 24.Энергетика живой клетки
Перечень вопросов, рассматриваемых в теме:
- Как энергия запасается в клетке;
- Что такое метаболизм;
- В чем суть процессов гликолиза, брожения и клеточного дыхания;
- Какие процессы проходят на световой и темновой фазах фотосинтеза;
- Как связаны процессы энергетического и пластического обмена;
- Что представляет собой хемосинтез.

Глоссарий по теме:
Метаболизм (обмен веществ) — сложная цепь превращений веществ в организме начиная с момента их поступления из внешней среды и кончая удалением продуктов распада. Представляет собой совокупность процессов энергетического обмена (катаболизма диссимиляции) и пластического обмена (анаболизма, ассимиляции).
Энергетический обмен – это совокупность химических реакций постепенного распада органических соединений, сопровождающихся высвобождением энергии, часть которой расходуется на синтез АТФ. Синтезированная АТФ становится универсальным источником энергии для жизнедеятельности организмов. Значение энергетического обмена – снабжение клетки энергией, которая необходима для жизнедеятельности.
Пластический обмен – это совокупность химических реакций образования (синтеза) из простых веществ с затратой энергии более сложные. Непосредственным поставщиком энергии в клетках выступает АТФ.
Фотосинтез – процесс образования органических веществ из неорганических (углекислого газа и воды) с использованием солнечной энергии.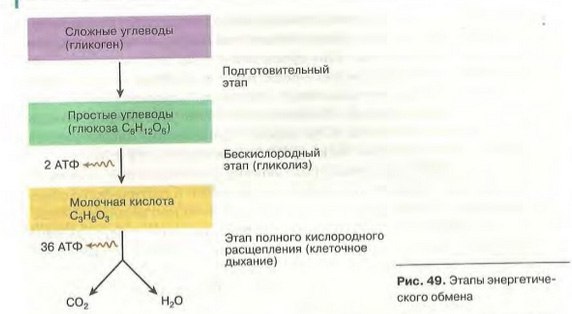 Проходит в два этапа: световая фаза (происходит улавливание и фиксация энергии света в АТФ) и темновая (связывание углекислого газа в молекулы глюкозы с затратой энергии АТФ).
Проходит в два этапа: световая фаза (происходит улавливание и фиксация энергии света в АТФ) и темновая (связывание углекислого газа в молекулы глюкозы с затратой энергии АТФ).
Хемосинтез — процесс образования органических веществ из неорганических (углекислого газа и воды) с использованием энергии окисления неорганических веществ. Например, такой тип питания используют азотфиксирующие бактерии.
Клеточное или тканевое дыхание — совокупность биохимических реакций, протекающих в клетках живых организмов, в ходе которых происходит окисление углеводов, липидов и аминокислот до углекислого газа и воды.
Основная и дополнительная литература по теме урока :
Естествознание. 10 класс [Текст]: учебник для общеобразоват. организаций: базовый уровень / И.Ю. Алексашина, К.В. Галактионов, И.С. Дмитриев, А.В. Ляпцев и др. / под ред. И.Ю. Алексашиной. – 3-е изд., испр. – М.: Просвещение, 2017.: с 115 — 118.
Электронные ресурсы:
Обмен веществ. Портал открытая биология // Электронный доступ: https://biology.ru/textbook/content.html
Портал открытая биология // Электронный доступ: https://biology.ru/textbook/content.html
АТФ и её роль в клетке .Проект «вся биология» // Электронный доступ: http://www.sbio.info/materials/obbiology/obbkletka/stroenorg/12
Энергетика живой клетки. Научно-познавательный журнал «Познавайка» // Электронный доступ: http://www.poznavayka.org/biologiya/energiya-zhivoy-kletki/
Энергетика живой клетки. Журнал «В МИРЕ НАУКИ» №3, 2006 . БИОЛОГИЯ // электронный доступ: https://elementy.ru/nauchno-populyarnaya_biblioteka/430308/430310?SSL=1
Теоретический материал для самостоятельного изучения
Обязательным условием существования биологических систем являются потоки энергии. В этом заключается ключевое различие между живой и неживой природой. Энергия не хранится в клетке, а поступает извне. Ключевую роль в трансформации энергии обеспечивает клетка, как элементарная структура живого. Специальные биохимические механизмы трансформируют одни виды энергии в другие, для обеспечения необходимых функций клетки.
Основным источником энергии для всех живых существ планеты Земля, является энергия Солнца. Однако эта энергия может быть использована живым только после того, как она будет усвоена фотоавтотрофами (от греч. «фото» — свет, «авто» — сам, «трофос» — питание).
В процессе эволюции появились и другие организмы, которые научились потреблять готовые органические соединения для получения запасённой в них энергии – гетеротрофы (от греч. «гетерос» — другой, «трофос» — питание).
Некоторые виды микроорганизмов (хемоавтотрофы) приобрели способность к использованию энергии, выделяемой при окислении неорганических веществ.
Таким образом, из всего многообразия существующих форм энергии живые существа на нашей планете используют только две – световую и энергию химических связей.
Главный переносчик энергии в клетке
Световая энергия Солнца и энергия, заключённая в потребляемой пище, запасаются в особых бимолекулярных аккумуляторах – молекулах АТФ (аденозинтрифосфат). В молекулах АТФ энергия запасается в виде высокоэнергетических химических связях между остатками фосфорной кислоты, которая освобождается при отщеплении фосфата: АТФ → АДФ + Ф + E.
В молекулах АТФ энергия запасается в виде высокоэнергетических химических связях между остатками фосфорной кислоты, которая освобождается при отщеплении фосфата: АТФ → АДФ + Ф + E.
Выделяемая энергия используется клетками для процессов выработки тепла, мышечных сокращений (мышечная клетка), для проведения нервного импульса (нервные клетки) и т.п.
Обратный процесс образования АТФ с затратой энергии, получил название энергетический обмен.
Синтез макромолекул важнейших органических соединений, необходимых для построения структур клетки, обеспечения всех процессов жизнедеятельности клеток – пластический обмен — обеспечивается также энергией АТФ.
Независимо от типа питания, универсальным аккумулятором энергии живых организмов выступают молекулы АТФ, где добытая энергия извне запасается в виде химических связей. Такая схожесть иллюстрирует единство происхождения всего живого.
Метаболизм
Поступившие вместе с пищей (или в результате фотосинтеза) органические вещества расщепляются на более простые (катаболизм или диссимиляция), которые служат для постройки макромолекул органических соединений (анаболизм или ассимиляция). Эти процессы происходят в организме одновременно. Совокупность этих процессов получила название – метаболизм. В результате его организм осуществляет обмен веществом и энергией с окружающей средой. Наибольшее значение для энергетического обмена являются многостадийные реакции расщепления глюкозы.
Эти процессы происходят в организме одновременно. Совокупность этих процессов получила название – метаболизм. В результате его организм осуществляет обмен веществом и энергией с окружающей средой. Наибольшее значение для энергетического обмена являются многостадийные реакции расщепления глюкозы.
На стадии гликолиза в цитоплазме клетки происходит ферментативное расщепление молекулы глюкозы с образованием более простой пировиноградной кислоты и молекул АТФ: С6Н12О6 + 2 АДФ + 2 Ф → 2С3Н4О3 + 4Н+ + 2АТФ
Молекулы пировиноградной кислоты обладают значительной энергией, высвобождение которой происходит в митохондриях. В ходе так называемого клеточного дыхания (аэробного расщепления), вещество распадается на углекислый газ, который впоследствии выделяется из клетки и воду. По последним исследованиям, при этом образуется 30 молекул АТФ.
Суммарную реакцию окисления глюкозы можно представить следующим образом:
С6Н12О6 + 6О2 + 6Н2О + 32 АДФ + 32 Ф → 6 СО2 + 12 Н2О + 32АТФ
Некоторые микроорганизмы при недостатке кислорода расщепляют глюкозу в процессе анаэробного дыхания или брожения.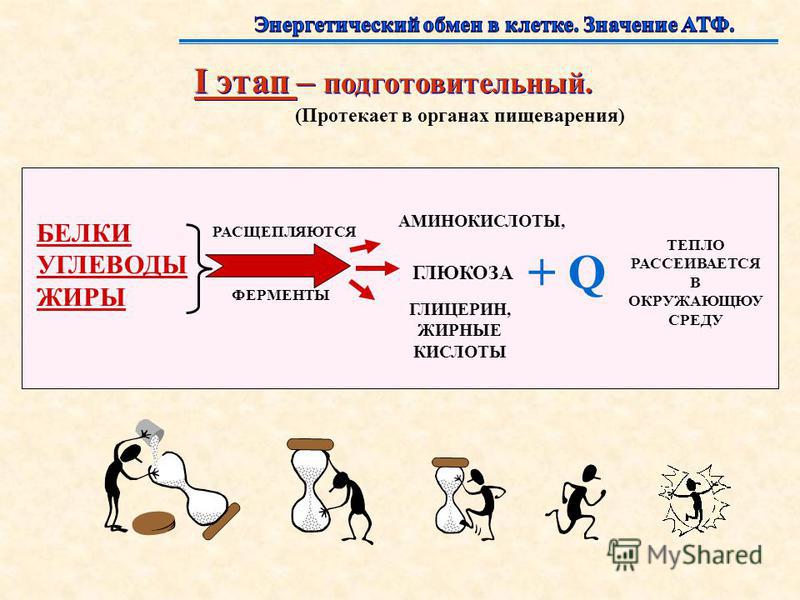 В зависимости от конечных продуктов такого расцепления различают спиртовое брожение (с образование этанола), молочнокислое (молочная кислота). Последнее происходит и в мышцах, при недостатке кислорода, например во время длительной тренировки. Энергетический выход такого типа расщепления менее энергоэффективен.
В зависимости от конечных продуктов такого расцепления различают спиртовое брожение (с образование этанола), молочнокислое (молочная кислота). Последнее происходит и в мышцах, при недостатке кислорода, например во время длительной тренировки. Энергетический выход такого типа расщепления менее энергоэффективен.
Основным источником энергии для организмов является окисление глюкозы в митохондриях. При этом также может происходить окисление других органических соединений (белков, жиров), потребляемых, например, вместе с пищей.
Фотосинтез
Фотоавтотрофы имеют уникальные ферментативные системы, способные трансформировать энергию солнечного света в энергию химической связи. Процесс образования органических веществ из неорганических (углекислого газа и воды) с использованием солнечной энергии получил название фотосинтез. В растениях фотосинтезирующие комплексы сосредоточены в специальных органеллах – хлоропластах. Основной пигмент – хлорофилл – выполняет функцию световых «антенн», улавливая световые волны практически всех диапазонов, кроме зелёного. Стоит отметить, что это обуславливает окраску листьев растений.
Стоит отметить, что это обуславливает окраску листьев растений.
В так называемой, световой фазе, кванты света выбивают электроны из молекулы хлорофилла, и он начинает передаваться по специальным белковым переносчикам, расположенных на мембране хлоропластов. Под действием света одновременно происходит разложение воды (фотолиз). В реакции высвобождается, в том числе катион водорода (Н+), необходимый для последующего биосинтеза, который захватывает молекула НАДФ (никотинамидадениндинуклеотидфосфат): НАДФ+ + Н+→НАДФ∙Н
Энергия возбуждённого электрона заряжает известный нам биологический катализатор АТФ и молекулу НАДФ – в этом заключается биологический смысл световой фазы фотосинтеза.
Заметим, что побочными продуктами фотолиза воды становятся свободный кислород и свободные электроны, восстанавливающие хлорофилл: 2Н2О→ Н+ + 4е— + О2
Дальнейший процесс может уже проходить без света. Сущность реакций темновой фазы можно выразить следующим уравнением: СО2 + НАДФ∙Н + АТФ = С6Н12О6 +АДФ + НАДФ+
Не сложно заметить, что выделяются вещества необходимые на начальном этапе фотосинтеза, что замыкает цикл.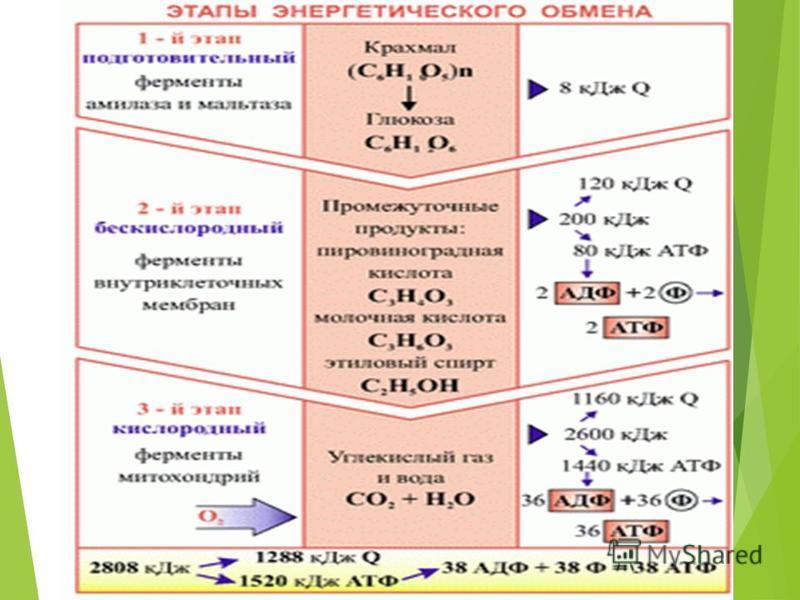 Энергия молекулярных аккумуляторов была использована для фиксации углекислого газа в энергию химических связей углевода.
Энергия молекулярных аккумуляторов была использована для фиксации углекислого газа в энергию химических связей углевода.
Фотосинтез, таким образом, является процессом превращения одной (световой) формы энергии в другую(химическую). Вся энергия биосферы запускается благодаря этому процессу. Другими словами, фотосинтез является отражением космических потоков энергии. Помимо этого, фотосинтезирующие организмы не только обеспечивают первичный синтез органических соединений, но и создают условия необходимые для существования других живых организмов.
Взаимосвязь энергетического и пластического обмена
Не сложно заметить, что процессы аккумулирования энергии в молекулах АТФ (энергетический обмен) и использование запасённой энергии для синтеза необходимых веществ (белков, жиров, углеводов, нуклеиновых кислот) неразрывно связаны. Так синтез АТФ не возможен без разложения органических веществ, а синтез веществ клетки не возможен без энергии АТФ. Причём, заметим, что и фотосинтез представляет собой единство этих процессов: темновая фаза – пластический обмен, световая фаза – энергетический.
Оба процесса протекают одновременно и неотделимы друг от друга, обеспечивая жизнедеятельность организма. Таким образом, в клетках происходит трансформация вещества и энергии, которые лежат в основе существования жизни и непрерывного самообновления. Сходство процессов энергетического обмена в клетках всех живых организмов является доказательством единства их происхождения.
Вывод
В клетках происходят одновременно процессы энергетического и пластического обмена, это лежит в основе сохранения жизни. Взаимообмен энергией и веществом между живой и неживой природой является иллюстрацией принципа единства и взаимосвязи материального мира.
Примеры и разбор решения заданий тренировочного модуля:
Задание 1. Выберите один ответ:
- Универсальным аккумулятором энергии в клетке является:
- Жиры;
- Белки;
- АТФ;
- НАДФ∙Н.
Ответ: АТФ
Пояснение: универсальной «разменной валютой» в энергетике живой клетки выступает АТФ. При его распаде выделяется энергия, которая расходуется на все жизненно важные процессы.
При его распаде выделяется энергия, которая расходуется на все жизненно важные процессы.
Задание 2. Исправьте ошибки, анализируя текст с позиции энергетического обмена:
В рационе питания человека помимо белков растительных и животных не обязательно должны присутствовать углеводы и жиры. Отсутствие жиров в пище не приводит к истощению. Человек толстеет, если употребляет в пищу избыточное количество углеводов. На сое и рисе можно прожить.
Ответ: В рационе питания человека помимо белков растительных и животных не обязательно должны присутствовать углеводы и жиры. Отсутствие жиров в пище не приводит к истощению. Человек толстеет, если употребляет в пищу много жиров. Исключительно на сое и рисе можно благополучно прожить.
Пояснение: с точки зрения энергетического обмена, наиболее энергоэффективными являются жиры. При этом, жиры, поступающие с пищей, используются в том числе, для построения многих важных соединений, например гормонов.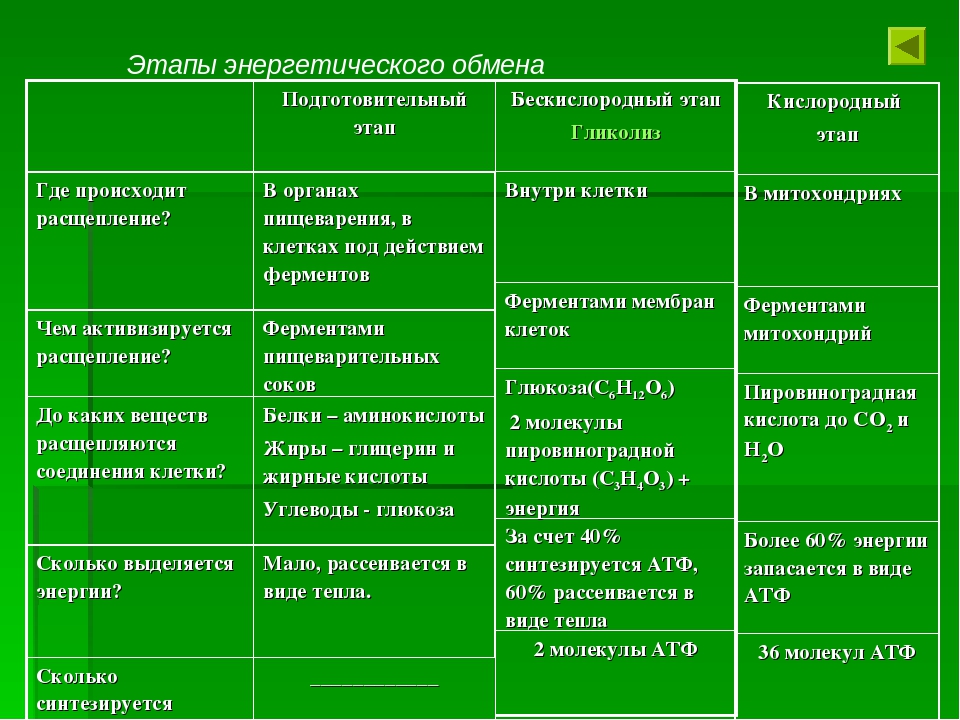 «Быстрая» энергия углеводов в избыточном количестве может приводить к полноте. Употребление только растительного белка в пищу, по сравнению с животным, является менее энергоэффективным и при отсутствии других источников энергии может приводить к истощению организма.
«Быстрая» энергия углеводов в избыточном количестве может приводить к полноте. Употребление только растительного белка в пищу, по сравнению с животным, является менее энергоэффективным и при отсутствии других источников энергии может приводить к истощению организма.
Урок 24. энергетика живой клетки — Естествознание — 10 класс
Энергетика живой клетки
Необходимо запомнить
ВАЖНО!
Обязательным условием существования биологических систем являются потоки энергии. В этом заключаются ключевые различия между живой и неживой природой. Ключевую роль в трансформации энергии обеспечивает клетка, как элементарная структура живого.
Из всего многообразия существующих форм энергии живые существа на нашей планете используют только две – световую и энергию химических связей. В зависимости от типа питания организмы разделают на автотрофов (от греч. «авто» — сам, «трофос» — питание) и гетеротрофов (от греч. «гетерос» — другой, «трофос» — питание) .
«гетерос» — другой, «трофос» — питание) .
Главным переносчиком энергии в клетке являются молекулы АТФ (аденозинтрифосфат) Энергия в АТФ запасается в виде высокоэнергетических химических связях между остатками фосфорной кислоты, которая освобождается при отщеплении фосфата:
АТФ → АДФ + Ф + E
Выделяемая энергия используется клетками для процессов выработки тепла, мышечных сокращений (мышечная клетка), для проведения нервного импульса (нервные клетки) и т.п.
Обратный процесс образования АТФ с затратой энергии, получил название энергетический обмен.
Синтез макромолекул важнейших органических соединений, необходимых для построения структур клетки, обеспечения всех процессов жизнедеятельности клеток – пластический обмен — обеспечивается также энергией АТФ.
Независимо от типа питания, универсальным аккумулятором энергии живых организмов выступают молекулы АТФ. Такая схожесть иллюстрирует единство происхождения всего живого.
Фотосинтез
Процесс образования органических веществ из неорганических (углекислого газа и воды) с использованием солнечной энергии получил название фотосинтез. Различают в фотосинтезе две фазы: световую и темовую.
Различают в фотосинтезе две фазы: световую и темовую.
В так называемой, световой фазе, кванты света выбивают электроны из молекулы хлорофилла ион начинает передаваться по специальным белковым переносчикам, расположенных на мембране хлоропластов. Под действием света одновременно происходит разложение воды (фотолиз). Энергия возбуждённого электрона используется на синтез АТФ и молекулу НАДФ (переносчик водорода) – в этом биологический смысл световой фазы фотосинтеза.
Побочными продуктами фотолиза воды становятся кислород и свободные электроны:
2Н2О→ Н+ + 4е— + О2
Сущность реакции темновой фазы можно выразить следующим уравнением:
СО2 + НАДФ∙Н + АТФ = С6Н12О6 +АДФ + НАДФ+
Фотосинтез, таким образом, является процессом превращения одной (световой) формы энергии в другую(химическую). Вся энергия биосферы запускается благодаря этому процессу. Другими словами, фотосинтез является отражением космических потоков энергии.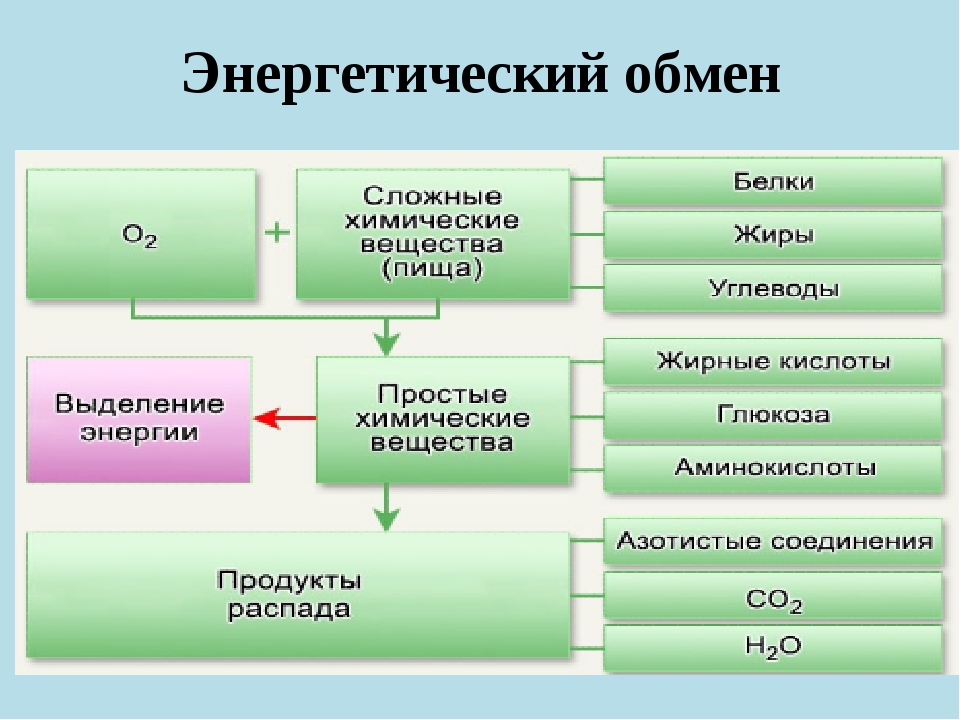 Помимо этого, фотосинтезирующие организмы не только обеспечивают первичный синтез органических соединений, но и создают условия необходимые для существования других живых организмов.
Помимо этого, фотосинтезирующие организмы не только обеспечивают первичный синтез органических соединений, но и создают условия необходимые для существования других живых организмов.
Метаболизм
Поступившие вместе с пищей (или в результате фотосинтеза) органические вещества расщепляются на более простые (катаболизм или диссимиляция), которые служат для построения макромолекул органических соединений (анаболизм или ассимиляция). Эти процессы происходят в организме одновременно. Совокупность этих процессов получила название – метаболизм. В результате его организм осуществляет обмен веществом и энергией с окружающей средой.
Наибольшее значение для энергетического обмена являются многостадийные реакции окисления глюкозы.
На стадии гликолиза (бескислородного расщепления) в цитоплазме клетки происходит ферментативное расщепление молекулы глюкозы с образованием более простой пировиноградной кислоты и молекул АТФ:
С6Н12О6 + 2 АДФ + 2 Ф → С3Н4О3 + 4Н+ + 2АТФ
Молекулы пировиноградной кислоты обладают значительной энергией, высвобождение которой происходит в митохондриях. В ходе так называемого клеточного дыхания, образуется 30 молекул АТФ.
В ходе так называемого клеточного дыхания, образуется 30 молекул АТФ.
Суммарную реакцию окисления глюкозы можно представить следующим образом:
С6Н12О6+6О2+6Н2О+32АДФ+32Ф→6СО2+12Н2О +32АТФ
Некоторые микроорганизмы при недостатке кислорода расщепляют глюкозу в процессе анаэробного дыхания или брожения. В зависимости от конечных продуктов такого расцепления различают спиртовое брожение (с образование этанола), молочнокислое (молочная кислота). Последнее происходит и в мышцах, при недостатке кислорода, например во время длительной тренировки. Энергетический выход такого типа расщепления менее энергоэффективен.
Основным источником энергии для организмов является окисление глюкозы в митохондриях. При этом также может происходить окисление других органических соединений (белков, жиров), потребляемых, например, вместе с пищей. Расщепление жиров происходит с более значительным выделением энергии (чем углеводов), но этот процесс более длительный. Потреблённые белки в первую очередь идут на построение собственных белков клетки, и вовлекаются в энергетический обмен в крайних случаях. Поэтому питание должно быть сбалансированным.
Потреблённые белки в первую очередь идут на построение собственных белков клетки, и вовлекаются в энергетический обмен в крайних случаях. Поэтому питание должно быть сбалансированным.
Взаимосвязь энергетического и пластического обмена
Процессы аккумулирования энергии в молекулах АТФ (энергетический обмен) и использование запасённой энергии для синтеза необходимых веществ (белков, жиров, углеводов, нуклеиновых кислот) неразрывно связаны. Так синтез АТФ не возможен без разложения органических веществ, а синтез веществ клетки не возможен без энергии АТФ.
Оба процесса протекают одновременно и неотделимы друг от друга, обеспечивая жизнедеятельность организма. Таким образом, в клетках происходит трансформация вещества и энергии, которые лежат в основе существования жизни и непрерывного самообновления. Сходство процессов энергетического обмена в клетках всех живых организмов является доказательством единства их происхождения.
Вывод
Добытая энергия извне запасается в универсальных биологических аккумуляторах АТФ в виде химических связей.
В клетках происходят одновременно процессы энергетического и пластического обмена, это лежит в основе сохранения жизни. Взаимообмен энергией и веществом между живой и неживой природой является иллюстрацией принципа единства и взаимосвязи материального мира.
Сравнение энергетического и пластического обменов
Механизм повреждения энергетического обмена при гипоксии и возможные пути его коррекции фумаратсодержащими растворами
Л.В. Слепнева, Г.А. Хмылова
ФГБУ «Российский НИИ гематологи трансфузиологии ФМБА», г. Санкт-Петербург
Трансфузиология №2, 2013
Резюме
Статья посвящена вопросам механизма действия препаратов, влияющих на процессы энергообразования в организме. Показаны пути коррекции нарушений энергетического обмена и преимущества фумаратсодержащих инфузионных растворов.
Ключевые слова: гипоксия, цикл Кребса, сукцинат, фумарат, фумаратсодержащие инфузионные растворы, мафусол, полиоксифумарин, конфумин.
В настоящее время нарушения энергетического обмена рассматриваются как один из ведущих патологических процессов, приводящих к необратимым последствиям и гибели организма, что обусловливает исключительную важность рассматриваемой проблемы. Коррекция или устранение энергодефицита является обязательным компонентом в лечении большинства патологических состояний, и в связи с этим, понимание механизма действия препаратов, способных влиять на различные звенья энергообмена, для практикующих врачей приобретают особую ценность.
Жизнедеятельность организма с многообразием всех физиологических функций и биохимических процессов возможна лишь при условии его постоянного энергообеспечения. В настоящее время имеется значительный экспериментальный и клинический материал, свидетельствующий о том, что различные экстремальные воздействия на организм (тяжелая кровопотеря, ожог, травма, сердечная недостаточность, острое отравление и др.) вызывают однотипные повреждения в клеточной системе энергообразования. Это явление обусловлено тем, что результирующим эффектом различных по своей природе экстремальных факторов является развитие острого кислородного голодания тканей. Дефицит кислорода — акцептора электронов в митохондриальной дыхательной цепи, приводит к глубокому подавлению биоэнергетической функции митохондрий. Выходит из строя основная энергетическая система клетки, энергопродукция клетками резко снижается, и, как следствие, нарушается течение многочисленных энергозависимых процессов в организме [2, 6, 11, 13, 24, 34, 36, 37, 43].
Это явление обусловлено тем, что результирующим эффектом различных по своей природе экстремальных факторов является развитие острого кислородного голодания тканей. Дефицит кислорода — акцептора электронов в митохондриальной дыхательной цепи, приводит к глубокому подавлению биоэнергетической функции митохондрий. Выходит из строя основная энергетическая система клетки, энергопродукция клетками резко снижается, и, как следствие, нарушается течение многочисленных энергозависимых процессов в организме [2, 6, 11, 13, 24, 34, 36, 37, 43].
Недостаточность систем энергообразования в клетке составляет существенный элемент патогенеза многих заболеваний. По мнению ряда авторов, поддержание жизни в экстремальных условиях возможно до тех пор, пока дефицит энергии не достигнет критических величин. Истощение клеточных энергетических резервов ниже допустимого уровня сопровождается развитием в клетке необратимых процессов и гибелью организма.
Прежде чем перейти к рассмотрению вопросов, связанных с нарушением энергетического обмена в клетке при патологических состояниях и его коррекции применением различных лекарственных средств, кратко остановимся на описании процессов энергообразования в нормально функционирующей клетке [16, 41].
На рис. 1 схематически представлен сложный процесс распада питательных веществ, который обеспечивает ступенчатое постепенное освобождение энергии и аккумуляцию ее в виде макроэргической фосфатной связи аденозинтрифосфата (АТФ).
Распад сложных питательных веществ на более простые является необходимым условием для дальнейшего использования их в клетке в качестве источников энергии и пластического материала. В катаболизме основных питательных веществ (углеводов, белков и жиров) можно выделить три основные стадии.
На первой стадии крупные молекулы под влиянием сложных ферментативных систем расщепляются на более простые. В результате действия этих ферментативных систем углеводы расщепляются до гексоз и пентоз, липиды – до глицерина и жирных кислот, из белков образуется около 20-ти аминокислот.
На второй стадии происходит дальнейшее расщепление образовавшихся соединений. Из 20-ти различных аминокислот образуется лишь несколько конечных продуктов, а именно, ацетил-коэнзим А, α-кетоглютаровая и щавелевоуксусная кислоты.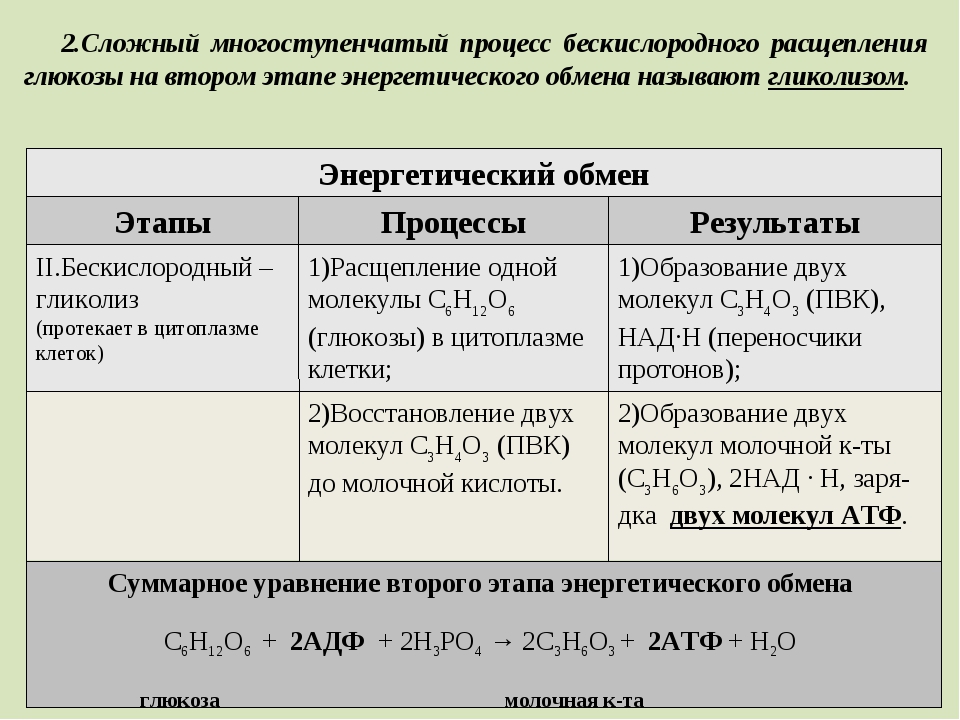
Жирные кислоты в процессе β-окисления превращаются в ацетил-КоА. Гексозы под действием ферментативных систем гликолиза расцепляются до пировиноградной кислоты, которая затем в процессе окислительного декарбоксилирования превращается также в ацетил-КоА.
Гликолиз является тем механизмом, посредством которого многие организмы получают химическую энергию из глюкозы и других субстратов в отсутствие молекулярного кислорода. У большинства аэробных организмов процесс гликолиза является предварительной ступенью для дальнейшего окисления продуктов брожения кислородом в процессе дыхания.
Метаболиты, образовавшиеся на второй стадии распада питательных веществ (ацетил-КоА, α-кето-глютаровая, щавелевоуксусная кислоты) вступают в третью стадию, которая для них является общей и на которой они в конечном итоге окисляются до СО2 и Н2О.
Третья стадия – стадия терминального окисления питательных веществ, во время которой освобождается основная масса энергии, осуществляется в митохондриях через цикл трикарбоновых кислот (ЦТК) и митохондриальную дыхательную цепь. ЦТК – общий конечный путь окислительного катаболизма всех видов клеточного топлива в аэробных условиях. В этом цикле под действием специфических дегидрогеназ протекают процессы дегидрирования субстратов, восстановительные эквиваленты от которых (протоны и электроны) поступают на митохондриальную дыхательную цепь. Дегидрирование – отщепление молекул Н2 от интермедиатов цикла Кребса происходит, в основном, при помощи дегидрогеназ, простетической группой которых является никотинамидаденин-динуклеотид (НАД), и лишь дегидрирование янтарной кислоты осуществляется ФАД-зависимой дегидрогеназой (сукцинатдегидрогеназой).
ЦТК – общий конечный путь окислительного катаболизма всех видов клеточного топлива в аэробных условиях. В этом цикле под действием специфических дегидрогеназ протекают процессы дегидрирования субстратов, восстановительные эквиваленты от которых (протоны и электроны) поступают на митохондриальную дыхательную цепь. Дегидрирование – отщепление молекул Н2 от интермедиатов цикла Кребса происходит, в основном, при помощи дегидрогеназ, простетической группой которых является никотинамидаденин-динуклеотид (НАД), и лишь дегидрирование янтарной кислоты осуществляется ФАД-зависимой дегидрогеназой (сукцинатдегидрогеназой).
Дыхательная цепь, состоящая из серии переносчиков электронов, передает восстановительные эквиваленты конечному акцептору электронов – молекулярному кислороду. Дыхательная цепь – это полиферментная система, локализованная во внутренней мембране митохондрий, основными компонентами которой являются НАД-зависимые дегидрогеназы, флавопротеиды и цитохромы (рис. 1).
Сопряженно с транспортом электронов протекает процесс окислительного фосфорилирования, в котором значительная часть свободной энергии электронов, передаваемых редокс-цепью на кислород, аккумулируется и трансформируется в специфическую макроэргическую связь АТФ.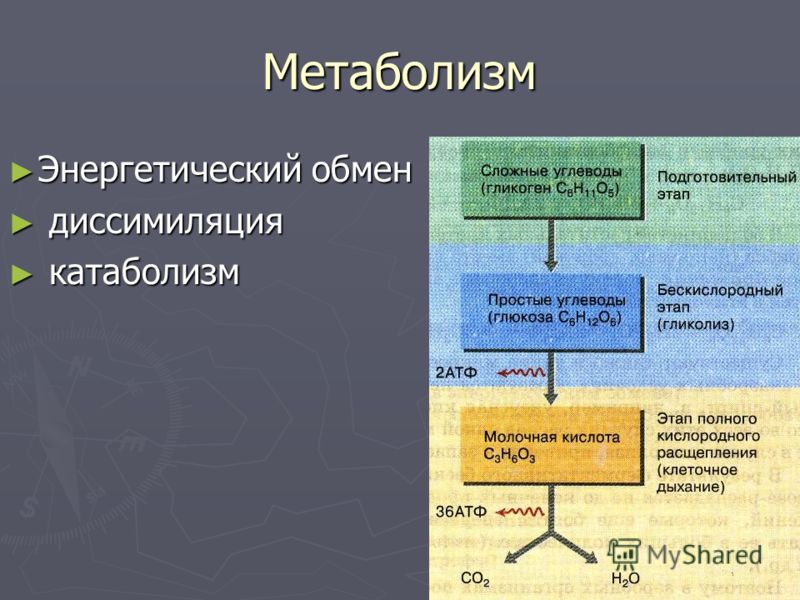 Таким образом, необходимая для нужд организма энергия образуется благодаря функционированию взаимосвязанных процессов гликолиза и дыхания. В процессе гликолиза высвобождается лишь незначительная часть той химической энергии, которая потенциально может быть извлечена из молекулы глюкозы. Полное окисление глюкозы до СО2 и Н 2О, осуществляемое в процессе дыхания, приводит к синтезу значительно большего количества макроэргов. При окислении одной молекулы глюкозы в гликолитическом цикле образуется 2 молекулы АТФ, тогда как дальнейшее расщепление продуктов гликолиза в цикле Кребса сопровождается синтезом 38 молекул АТФ. Таким образом, митохондриальная дыхательная цепь является основным местом приложения и утилизации кислорода в клетке.
Таким образом, необходимая для нужд организма энергия образуется благодаря функционированию взаимосвязанных процессов гликолиза и дыхания. В процессе гликолиза высвобождается лишь незначительная часть той химической энергии, которая потенциально может быть извлечена из молекулы глюкозы. Полное окисление глюкозы до СО2 и Н 2О, осуществляемое в процессе дыхания, приводит к синтезу значительно большего количества макроэргов. При окислении одной молекулы глюкозы в гликолитическом цикле образуется 2 молекулы АТФ, тогда как дальнейшее расщепление продуктов гликолиза в цикле Кребса сопровождается синтезом 38 молекул АТФ. Таким образом, митохондриальная дыхательная цепь является основным местом приложения и утилизации кислорода в клетке.
При дефиците кислорода – конечного акцептора электронов в редокс-цепи митохондрий – отмечается выраженная гиперредукция всех компонентов терминального звена окисления. Прекращаются транспорт электронов по дыхательной цепи и сопряженный с ним процесс образования макроэнергических фосфатов.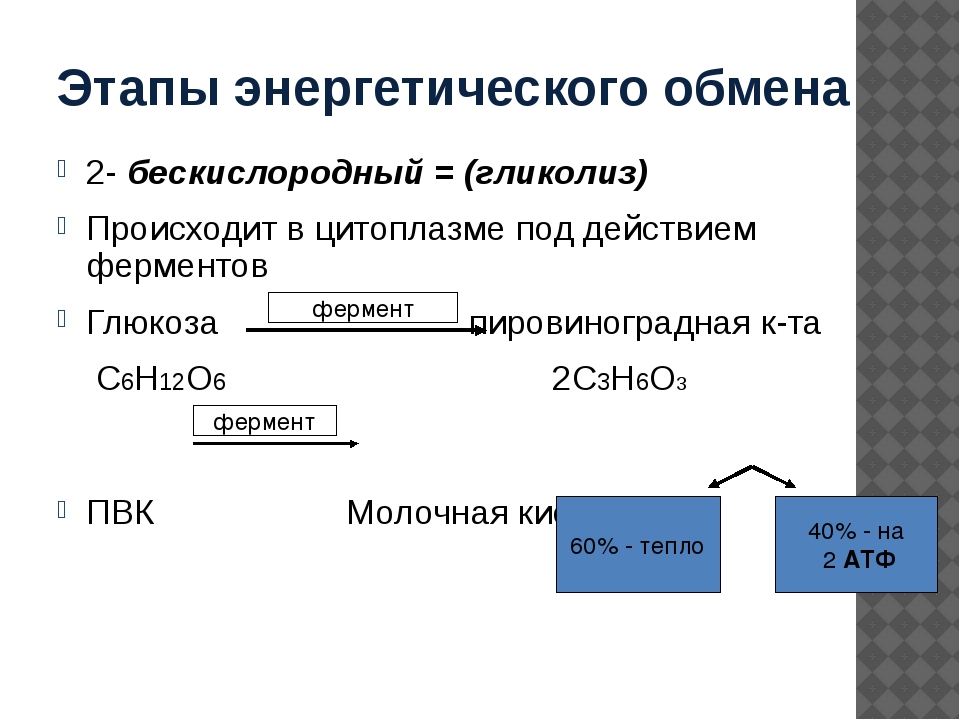 Известно, что в условиях нормоксии НАД-звено дыхательной цепи митохондрий принимает восстановительные эквиваленты из различных источников:
Известно, что в условиях нормоксии НАД-звено дыхательной цепи митохондрий принимает восстановительные эквиваленты из различных источников:
1. от субстратов цикла Кребса при участии специфических дегидрогеназ;
2. оксиацил-КоА-дегидрогеназы поставляютионы водорода на НАД- звено при окислении жирных кислот;
3. сложная система пируват-дегидрогеназы, отщепляя ионы водорода в реакциях окислительного декарбоксилирования, передает их на НАД-звено редокс-цепи;
4. внемитохондриальный НАД-Н, образованный в центральной реакции гликолитической оксидоредукции, также отдает свои протоны на митохондриальную дыхательную цепь (рис.1).
Мощный поток восстановительных эквивалентов в условиях кислородной недостаточности не может реализоваться из-за гипервосстановленности НАД-зависимого участка дыхательной цепи. Выключается из функционирования основная энергетическая система клетки, резко снижается продукция АТФ.
В анаэробных условиях клетка стремится восполнить энергетический дефицит за счёт активации гликолиза.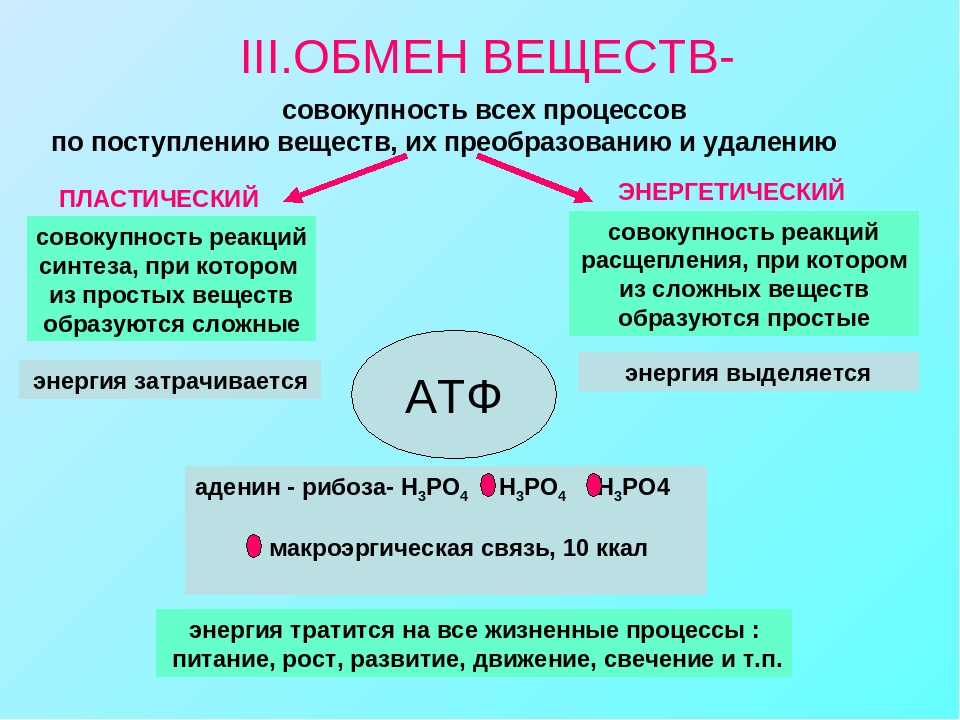 Несмотря на то, что при анаэробном гликолизе продуцируется почти в 20 раз меньше АТФ, чем при полном сгорании глюкозы в цикле Кребса, потенциальная скорость процесса в основном может обеспечить энергозатраты организма. Однако для осуществления гликолитических реакций необходим постоянный приток окисленной формы НАД, который при нормоксии обеспечивается работой специфических челночных механизмов. Гликолитический НАД-Н проникает через митохондриальные мембраны посредством функционирования α-глицерофосфатного, β-оксибутиратного и других механизмов переноса восстановительных эквивалентов.
Несмотря на то, что при анаэробном гликолизе продуцируется почти в 20 раз меньше АТФ, чем при полном сгорании глюкозы в цикле Кребса, потенциальная скорость процесса в основном может обеспечить энергозатраты организма. Однако для осуществления гликолитических реакций необходим постоянный приток окисленной формы НАД, который при нормоксии обеспечивается работой специфических челночных механизмов. Гликолитический НАД-Н проникает через митохондриальные мембраны посредством функционирования α-глицерофосфатного, β-оксибутиратного и других механизмов переноса восстановительных эквивалентов.
Оксибутиратный и глицерофосфатные шунты являются основными конкурентами лактатдегидрогеназного механизма окисления гликолитического НАД-Н, деятельность которого при нормальной концентрации кислорода в клетке подавлена более активными вышеназванными механизмами.
При нарушении электронтранспортной функции редокс-цепи и гиперредукции ее НАД-звена клетка вынуждена изыскивать другие пути реокисления цитоплазматического НАД-Н.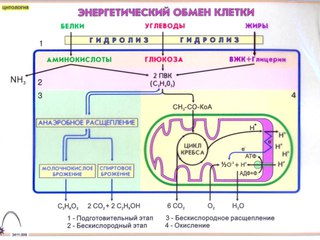 В условиях острой гипоксии конечный продукт гликолиза – пируват – не подвергается декарбоксилированию и не вовлекается в цикл Кребса, а, принимая восстановительные эквиваленты от цитоплазматического НАД-Н, превращается в лактат с освобождением новых порций окисленной формы НАД (рис. 2).
В условиях острой гипоксии конечный продукт гликолиза – пируват – не подвергается декарбоксилированию и не вовлекается в цикл Кребса, а, принимая восстановительные эквиваленты от цитоплазматического НАД-Н, превращается в лактат с освобождением новых порций окисленной формы НАД (рис. 2).
Активация лактатдегидрогеназного механизма поставки НАД для гликолиза в конечном итоге приводит к истощению запасов гликогена и тканевому ацидозу вследствие накопления кислых продуктов метаболизма (лактата, пирувата, оксибутирата, глицерофосфата и др.). Избыточные концентрации конечного субстрата анаэробного гликолиза – лактата – тормозят последнюю реакцию гликолитического цикла.
Регенерация НАД прекращается, и, именно, дефицит пиридиннуклеотида останавливает гликолиз и анаэробную продукцию АТФ. Клеточный ацидоз способствует нарушению проницаемости мембран, вплоть до разрушения лизосом. В цитоплазму поступают аутолитические ферменты. Развивается процесс аутолиза клеток, сопровождающийся повреждением тканей и органов.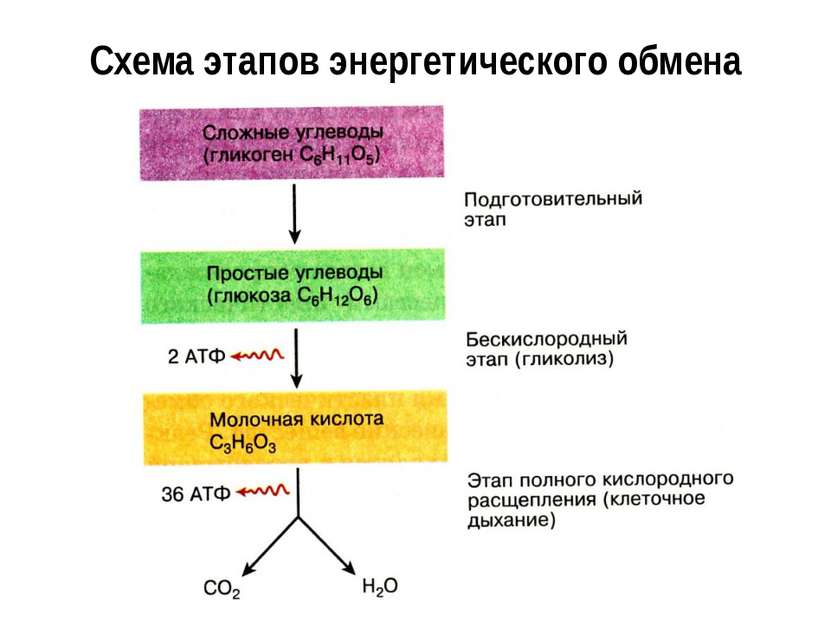 В организме формируются необратимые изменения.
В организме формируются необратимые изменения.
Таким образом, степень повреждения митохондриального метаболизма в условиях тяжелой кислородной недостаточности определяет тяжесть многих патологических состояний. Накопленный опыт лечения шока и кровопотери показывает, что существующие инфузионнотрансфузионные среды, проявляя лечебное действие в стадиях легкой и средней тяжести, оказываются недостаточными на поздних стадиях процесса. Особенности течения поздних стадий геморрагического шока связывают главным образом с генерализованными нарушениями метаболизма и возникающими в результате этого расстройствами энергообмена.
В связи с этим применение совместно с кровезаменителями препаратов, способных повысить энергетический потенциал клетки в условиях гипоксии, рассматривается как один из путей повышения эффективности инфузионной терапии гиповолемических состояний.
В ликвидации энергетического дефицита большое значение придается антигипоксантам. К настоящему времени не выработано единого общепринятого определения антигипоксантов и их классификации, так как в ответ на гипоксическое воздействие вовлекаются самые разные системы организма. Препараты биоэнергетического действия можно разделить на несколько групп.
Препараты биоэнергетического действия можно разделить на несколько групп.
К первой группе следует отнести препараты, являющиеся источником энергетического сырья (глюкоза, сорбит, АТФ, фосфорилированные гексозы и др.). Использование их показано при патологических состояниях, сопровождающихся истощением энергетических ресурсов в клетке. Включение в состав противошоковых кровезаменителей 5-10% глюкозы или фосфорилированных гексоз [1, 42] для поддержания гликолиза в клетках не позволяет существенно повысить эффективность инфузионной терапии из-за неизбежно возникающего накопления кислых продуктов метаболизма и дефицита окисленной формы пиридиннуклеотида (НАД). Отсюда понятно, что введение таких субстратов окисления, как глюкоза или гексозы, при гипоксии целесообразно лишь с препаратами, ускоряющими утилизацию лактата. Таким свойством обладают соединения группы гутимина. В эксперименте показан антигипоксический эффект гутимина и амтизола при геморрагическом шоке [8].
Ко второй группе препаратов можно отнести средства, которые, не являясь энергетически богатыми соединениями, способны активно воздействовать на энергетический обмен посредством коррекции отдельных звеньев многоступенчатого процесса аккумуляции энергии в клетке. Данные о нарушении транспорта электронов в дыхательной цепи митохондрий при шоке и кровопотери [28] являются теоретической предпосылкой для применения антигипоксантов с электрон-акцепторными свойствами. В литературе имеются довольно обширные сведения о применении естественных и искусственных антигипоксантов – переносчиков электронов. К числу первых относится цитохром С, который, как известно, является одним из компонентов дыхательной цепи митохондрий и служит мобильным переносчиком электронов. Играя важную роль в энергетическом метаболизме клетки, цитохром С показал высокую лечебную эффективность в клинической практике при терапии шока, кровопотери и постишемической гипоксии [12, 32].
Данные о нарушении транспорта электронов в дыхательной цепи митохондрий при шоке и кровопотери [28] являются теоретической предпосылкой для применения антигипоксантов с электрон-акцепторными свойствами. В литературе имеются довольно обширные сведения о применении естественных и искусственных антигипоксантов – переносчиков электронов. К числу первых относится цитохром С, который, как известно, является одним из компонентов дыхательной цепи митохондрий и служит мобильным переносчиком электронов. Играя важную роль в энергетическом метаболизме клетки, цитохром С показал высокую лечебную эффективность в клинической практике при терапии шока, кровопотери и постишемической гипоксии [12, 32].
Разработке и исследованию искусственных переносчиков электронов посвящено значительное количество работ. Эти соединения способны модифицировать дыхательную цепь митохондрий так, чтобы осуществлять «сброс» восстановительных эквивалентов непосредственно на кислород, минуя заблокированные участки дыхательной цепи. К числу таких веществ относится ряд соединений из класса хинонов (ортопарабензохиноны, нафтохиноны, гексогидрохиноны). Высокий редокс-потенциал этих препаратов определяет их способность к транспорту электронов [30]. При проведении экспериментов на животных многие из этих соединений оказались токсичными, что не позволило рекомендовать их в качестве лечебных средств. Из всех средств, формирующих искус- ственные редокс-системы, в медицинскую практику внедрен препарат «Гипоксен», представляющий собой синтетический полихинон [9].
К числу таких веществ относится ряд соединений из класса хинонов (ортопарабензохиноны, нафтохиноны, гексогидрохиноны). Высокий редокс-потенциал этих препаратов определяет их способность к транспорту электронов [30]. При проведении экспериментов на животных многие из этих соединений оказались токсичными, что не позволило рекомендовать их в качестве лечебных средств. Из всех средств, формирующих искус- ственные редокс-системы, в медицинскую практику внедрен препарат «Гипоксен», представляющий собой синтетический полихинон [9].
Известно, что антигипоксанты группы хинонов осуществляют перенос электронов с НАД-Н звена на кислород, минуя все 3 пункта фосфорилирования в дыхательной цепи и, следовательно, устранение дефицита энергии при введении этих препаратов может происходить лишь за счет активации гликолитической выработки АТФ. Однако для полноценного проявления антигипоксических свойств этих соединений необходим акцептор электронов – кислород. Наибольший интерес для включения в состав новых комплексных кровезаменителей представляют, так называемые, истинные антигипоксанты или антигипоксанты прямого действия, непосредственно влияющие на митохондриальный метаболизм при гипоксии.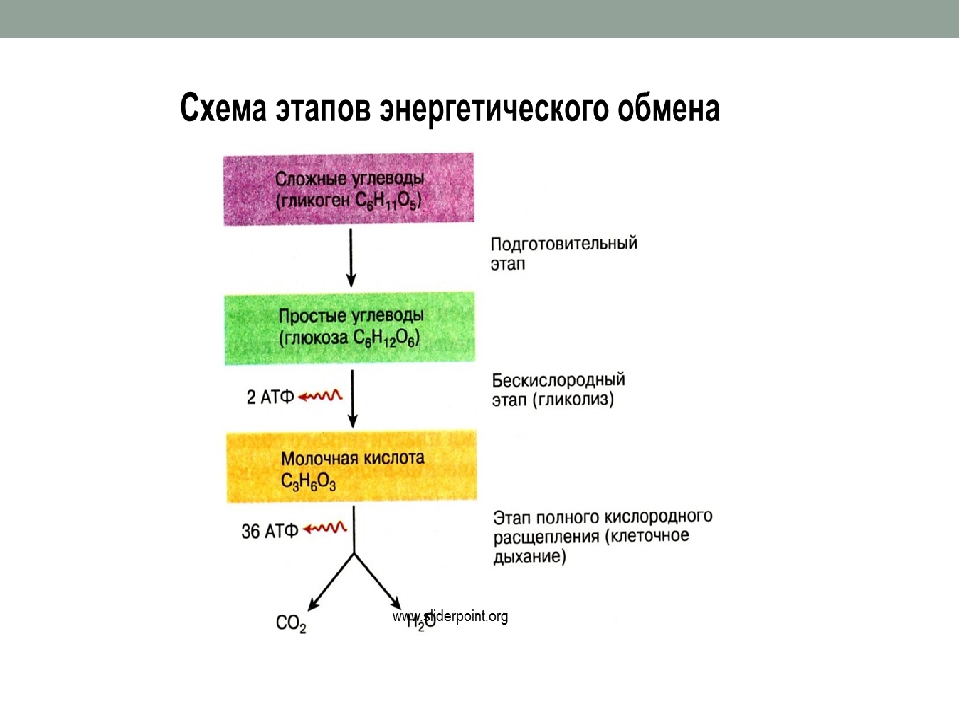
Согласно теоретическим предпосылкам, одна из возможностей поддержания биоэнергетики клетки может быть реализована посредством стимуляции адаптационных механизмов к гипоксии, работающих на последних этапах цикла Кребса. Из всех субстратов цикла Кребса только влияние сукцината на энергетический обмен подробно изучено в эксперименте на животных.
Исследование механизма действия сукцината при гипоксических состояниях проведено в работах М.Н. Кондрашовой с соавторами [5, 6]. Исследователи считают, что в условиях гипоксии сукцинат, не являясь НАД-зависимым субстратом, «монополизирует» дыхательную цепь и активно в ней окисляется. Высокая скорость реакции окисления сукцината, поставляющей 2 молекулы АТФ, позволяет компенсировать выработку 3-х молекул АТФ, образующихся при окислении НАД-зависимых субстратов. Однако в условиях прогрессирующей гипоксии дефицит кислорода, лимитирующий скорость окисления всех субстратов, снижает ценность сукцината и ставит его в один ряд с другими субстратами окисления.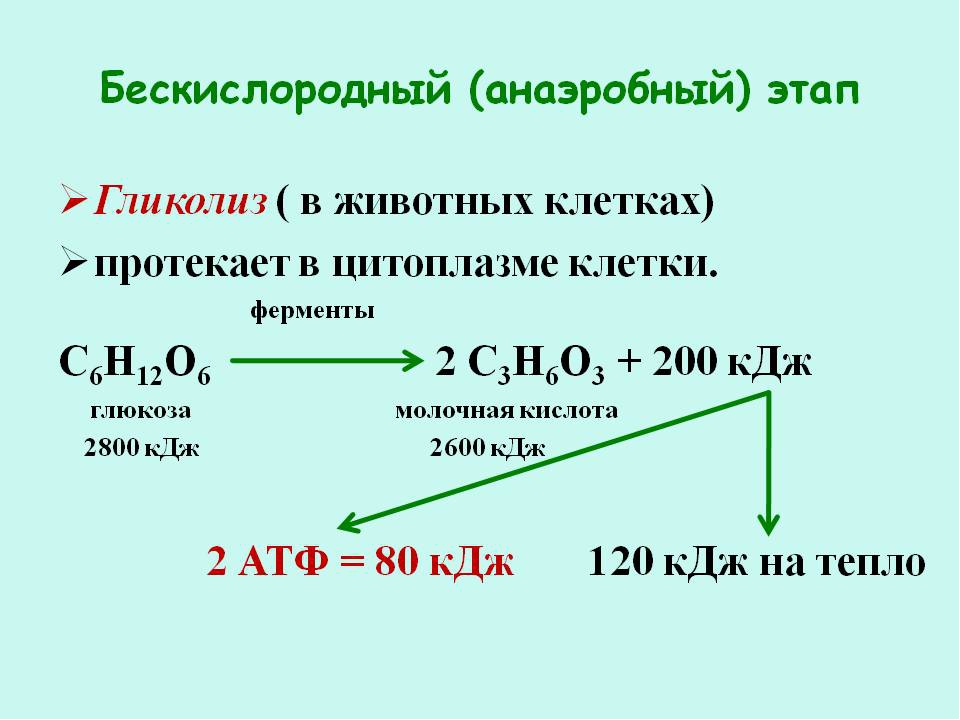 Поэтому применение сукцината в качестве антигипоксанта должно быть особенно эффективно в комплексе с препаратами, улучшающими кислородообеспечение организма. Преимущественное использование сукцината – естественная защита клетки против гипоксии. При этом пополнение фонда субстрата может происходить за счет реакций цикла Кребса, идущих как в прямом, так и в обратном направлениях (рис. 3).
Поэтому применение сукцината в качестве антигипоксанта должно быть особенно эффективно в комплексе с препаратами, улучшающими кислородообеспечение организма. Преимущественное использование сукцината – естественная защита клетки против гипоксии. При этом пополнение фонда субстрата может происходить за счет реакций цикла Кребса, идущих как в прямом, так и в обратном направлениях (рис. 3).
При обратном течении реакций имеющийся запас малата по мере необходимости превращается в фумарат, который восстанавливается в сукцинат. Восстановление фумарата сопровождается выработкой АТФ, и поэтому реакции обращения в системе «малат-фумарат-сукцинат» способны поддерживать окислительное фосфорилирование даже при аноксии.
В условиях же гипоксии инверсивные превращения фумарата выполняют роль триггера, который, в зависимости от концентрации кислорода регулирует течение конечных реакций цикла Кребса в прямом либо в обратном направлениях, и эти реакции сопровождаются синтезом АТФ. Механизм инверсивных превращений фумарата в цикле Кребса объясняет эффективность применения фумаратсодержащих инфузионных сред, таких как кристаллоидный раствор – мафусол, коллоидный кровезаменитель – полиоксифумарин и концентрированный раствор фумарата натрия – конфумин. Эти препараты разработаны и основательно изучены в Российском НИИ гематологии и трансфузиологии. Лечебная эффективность была изучена на моделях геморрагического и ожогового шока, а также при экспериментальном перитоните [17–20, 23, 25,45]. Оценку эффективности инфузионных растворов определяли по совокупности показателей системной гемодинамики, кислородного режима, кислотно-основного состояния (КОС), перекисного окисления липидов и митохондриального метаболизма в печени и сердце животных. Полярографическое исследование митохондрий, выделенных из печени и сердца животных, леченных фумаратсодержащими растворами, свидетельствовало о полном восстановлении энергопродуцирующих функций этих органелл. Следует отметить, что летальность животных в контрольной группе (тяжелый шок) составляло 100%, при лечении мафусолом или полиоксифумарином – 17–20%.
Эти препараты разработаны и основательно изучены в Российском НИИ гематологии и трансфузиологии. Лечебная эффективность была изучена на моделях геморрагического и ожогового шока, а также при экспериментальном перитоните [17–20, 23, 25,45]. Оценку эффективности инфузионных растворов определяли по совокупности показателей системной гемодинамики, кислородного режима, кислотно-основного состояния (КОС), перекисного окисления липидов и митохондриального метаболизма в печени и сердце животных. Полярографическое исследование митохондрий, выделенных из печени и сердца животных, леченных фумаратсодержащими растворами, свидетельствовало о полном восстановлении энергопродуцирующих функций этих органелл. Следует отметить, что летальность животных в контрольной группе (тяжелый шок) составляло 100%, при лечении мафусолом или полиоксифумарином – 17–20%.
Результаты исследования митохондриального метаболизма позволяют предположить, что парентеральное введение фумарата индуцирует суперкомпенсацию адаптационного механизма к гипоксии, функционирующих на последних этапах цикла Кребса.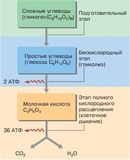 Фумарат в системе «малат-фумарат-сукцинат» способен поддерживать синтез АТФ как в аэробных, так и в анаэробных условиях. При дефиците кислорода фумарат, восстанавливаясь ФАД∙Н2-группой сукцинатдегидрогеазой, превращается в сукцинат и освобождает новые порции окисленной формы ФАД. Принимая восстановительные эквиваленты от НАД-Н, ФАД способствует снятию гипервосстановленности НАД звена дыхательной цепи и синтезу АТФ в бескислородной среде. При поступлении кислорода в клетку сукцинат, синтезируемый из фумарата, монополизирует дыхательную цепь и, активно окисляясь в ней, продуцирует АТФ (рис. 3). К тому же, образование в этих реакциях окисленной формы НАД запускает также и механизм гликолитической продукции АТФ. Поддержание энергетического потенциала клетки при инфузия фумарата способствует удлинению периода обратимых изменений в организме и предотвращает развитие «необратимости» при патологических состояниях, отягощенных глубокой гипоксией.
Фумарат в системе «малат-фумарат-сукцинат» способен поддерживать синтез АТФ как в аэробных, так и в анаэробных условиях. При дефиците кислорода фумарат, восстанавливаясь ФАД∙Н2-группой сукцинатдегидрогеазой, превращается в сукцинат и освобождает новые порции окисленной формы ФАД. Принимая восстановительные эквиваленты от НАД-Н, ФАД способствует снятию гипервосстановленности НАД звена дыхательной цепи и синтезу АТФ в бескислородной среде. При поступлении кислорода в клетку сукцинат, синтезируемый из фумарата, монополизирует дыхательную цепь и, активно окисляясь в ней, продуцирует АТФ (рис. 3). К тому же, образование в этих реакциях окисленной формы НАД запускает также и механизм гликолитической продукции АТФ. Поддержание энергетического потенциала клетки при инфузия фумарата способствует удлинению периода обратимых изменений в организме и предотвращает развитие «необратимости» при патологических состояниях, отягощенных глубокой гипоксией.
Парентеральное введение фумаратсодержащих растворов наряду с восстановлением биоэнергетики клетки, сопровождается «мягким» ощелачивающим действием препаратов на кислотно-основное состояние крови при ацидозе. Это действие обусловлено тем, что такие органические соли, как фумарат-, ацетат-, лактат-, сукцинат- и малат натрия являются соединениями, образованными сильным основанием (NaOH) и слабой кислотой. При гидролизе подобных солей в кровеносном русле освобождается соответствующая кислота и NaOH, который расходуется на нейтрализацию кислых продуктов метаболизма. Реакция гидролиза смещена вправо, так как постоянно происходит потребление продукта гидролитической реакции – NaOH (рис. 4).
Это действие обусловлено тем, что такие органические соли, как фумарат-, ацетат-, лактат-, сукцинат- и малат натрия являются соединениями, образованными сильным основанием (NaOH) и слабой кислотой. При гидролизе подобных солей в кровеносном русле освобождается соответствующая кислота и NaOH, который расходуется на нейтрализацию кислых продуктов метаболизма. Реакция гидролиза смещена вправо, так как постоянно происходит потребление продукта гидролитической реакции – NaOH (рис. 4).
Следует отметить, что вышеназванные соли оказывают мягкое ощелачивающие действие по сравнению с бикарбонатом натрия, широко используемым в клинической практике для ликвидации ацидоза. Реакция гидролиза NaHСО3 протекает значительно быстрее, так как в ходе реакции удаляются оба ее продукта: NaOH расходуется на нейтрализацию метаболитов, а второй продукт реакции – угольная кислота, нестоек и разлагается на Н2О и СО2. Образованная в избыточном количестве щелочь может способствовать развитию алкалоза, что имеет место в клинических условиях при передозировке бикарбоната натрия.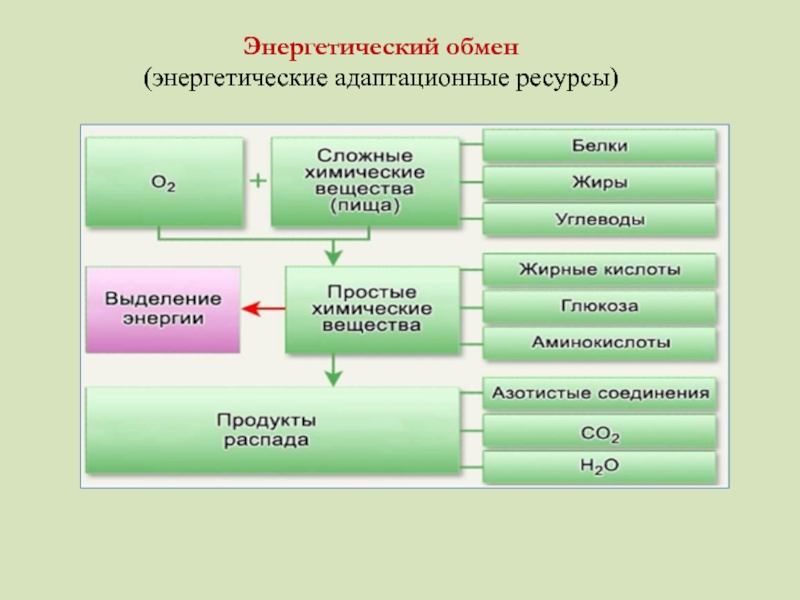
Все вышеперечисленные соли входят в состав различных инфузионных растворов (мафусол, полиоксифумарин, конфумин, лактасол, Рингер-лактат, ацесоль, реамберин, стерофундин и др.). Однако оказывая ощелачивающее действие при ацидозе, далеко не все эти препараты способны поддержать энергетический обмен при гипоксии. Восстановление показателей КОС «химическим путем» является недостаточным для успешной терапии шока.
Следует к тому же учитывать, что при гидролизе лактата натрия выделяется молочная кислота, которая в сумме с эндогенной молочной кислотой, возникающей в больших концентрациях при гипоксии, могут способствовать подавлению реакций гликолиза, что, в свою очередь, вызывает снижение продукции гликолитической АТФ. Существуют также исследования, указывающие, что лактат может вызвать интерстициальный отек головного мозга и повышать агрегацию тромбоцитов и эритроцитов [14, 39, 44]. Лактатсодержащие инфузионные растворы нельзя использовать при печеночной недостаточности [35, 38, 40], а также в случаях шока, сопровождающегося гиперлактатемией или лактатным ацидозом [33].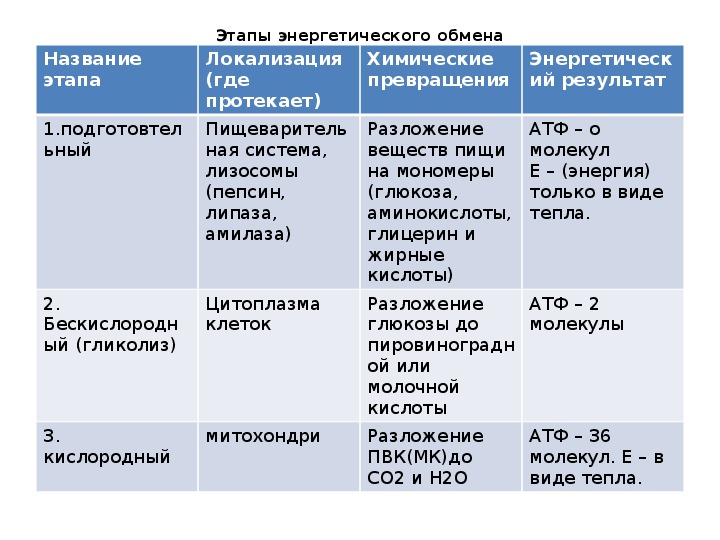
Ацетат натрия, в отличие от лактата, не проявляет токсического действия при тяжелом шоке. Однако утилизация уксусной кислоты, образованной при гидролизе ацетата натрия, в условиях кислородной недостаточности затруднена вследствие постгипоксического дефекта в функционировании митохондриальной дыхательной цепи. Лечебное действие фумарата натрия в сравнении с лактатом и ацетатом представляется более физиологичным, так как при его введении наряду с ощелачивающим эффектом проявляется и его влияние на восстановление процессов генерации энергии в митохондриях, а, следовательно, устраняется причина возникновения метаболического ацидоза.
Сукцинатсодержащие растворы, в частности «Реамберин», способствуют поддержанию энергетического обмена, однако, в условиях острого дефицита кислорода подавляется окисление сукцината и существенно снижается его энергопродуцирующая функция. Окисление малата в цикле Кребса осуществляется НАД-зависимой малатдегидрогеназой, и эта реакция тормозится из-за гипервосстановленности НАД-звена редокс-цепи митохондрий при гипоксии.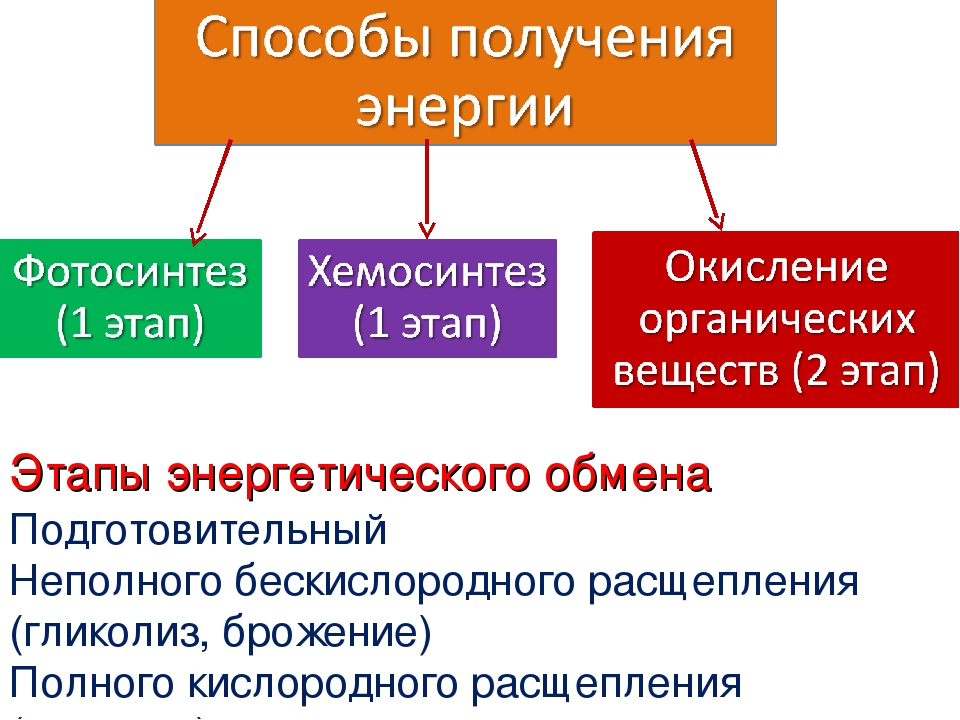 Следовательно, в этих условиях субстрат не способен повысить энергетический потенциал клетки. К тому же, в инфузионном малатсодержащем растворе «Стерофундин» концентрация малата очень низкая, чтобы обеспечить достаточную продукцию АТФ. В условиях гипоксии повышение концентрации малата могло бы создать условия для обращения реакций в цикле Кребса с увеличением фонда фумарата, способного принимать восстановительные эквиваленты (Н2) и синтезировать АТФ. Однако концентрация малата в стерофундине (5 ммоль/л) незначительна для запуска реакций в цикле Кребса в обратном направлении.
Следовательно, в этих условиях субстрат не способен повысить энергетический потенциал клетки. К тому же, в инфузионном малатсодержащем растворе «Стерофундин» концентрация малата очень низкая, чтобы обеспечить достаточную продукцию АТФ. В условиях гипоксии повышение концентрации малата могло бы создать условия для обращения реакций в цикле Кребса с увеличением фонда фумарата, способного принимать восстановительные эквиваленты (Н2) и синтезировать АТФ. Однако концентрация малата в стерофундине (5 ммоль/л) незначительна для запуска реакций в цикле Кребса в обратном направлении.
Фумаратсодержащие растворы (мафусол, полиоксифумарин) содержат высокие концентрации фумарата (86 ммоль/л), обеспечивающие как выработку АТФ, так и накопление сукцината, который активно окисляется при поступлении кислорода. Введение субстратов в организм при гипоксии показано еще и вследствие того, что кислородная недостаточность сопровождается значительным субстратным голодом клетки. Препараты «Мафусол» и «Полиоксифумарин» с высокой концентрацией фумарата и возможностью инфузий больших объемов этих растворов без побочных эффектов являются высокоэффективными средствами терапии шока различного генеза. Это подтверждено клинически. Так, кристаллоидный кровезаменитель «Мафусол» разрешен к медицинскому применению уже более 20 лет и широко используется в разных областях медицины (хирургия, неврология, кардиология, реаниматология, педиатрия, акушерство и гинекология, комбустиология, токсикология и др.) [3, 15, 22, 27, 29, 31]. Отличительной особенностью этого препарата является то, что его можно переливать в больших количествах, не только внутривенно, но и внутриартериально, а также в смеси для заполнения контура АИК при открытых операциях на сердце. Ни один из существующих сейчас на фармацевтическом рынке инфузионных антигипоксических препаратов не обладает этими свойствами. Полифункциональный коллоидный плазмозаменитель «Полиоксифумарин» с 1999 года успешно применяется у взрослых и детей в клинической практике гиповолемических состояний различной степени тяжести [10, 21, 22]. Аналогов ему нет ни в России, ни зарубежом.
Это подтверждено клинически. Так, кристаллоидный кровезаменитель «Мафусол» разрешен к медицинскому применению уже более 20 лет и широко используется в разных областях медицины (хирургия, неврология, кардиология, реаниматология, педиатрия, акушерство и гинекология, комбустиология, токсикология и др.) [3, 15, 22, 27, 29, 31]. Отличительной особенностью этого препарата является то, что его можно переливать в больших количествах, не только внутривенно, но и внутриартериально, а также в смеси для заполнения контура АИК при открытых операциях на сердце. Ни один из существующих сейчас на фармацевтическом рынке инфузионных антигипоксических препаратов не обладает этими свойствами. Полифункциональный коллоидный плазмозаменитель «Полиоксифумарин» с 1999 года успешно применяется у взрослых и детей в клинической практике гиповолемических состояний различной степени тяжести [10, 21, 22]. Аналогов ему нет ни в России, ни зарубежом.
Применение концентрированного раствора фумарата натрия (препарата «Конфумин») в качестве антигипоксического компонента в схемах инфузионно-трансфузионной терапии существенно увеличивает уровень субстратов окисления в кровеносном русле и позволяет повысить лечебную эффективность общепринятых в клинической практике плазмозаменителей [3, 4, 22, 25-27].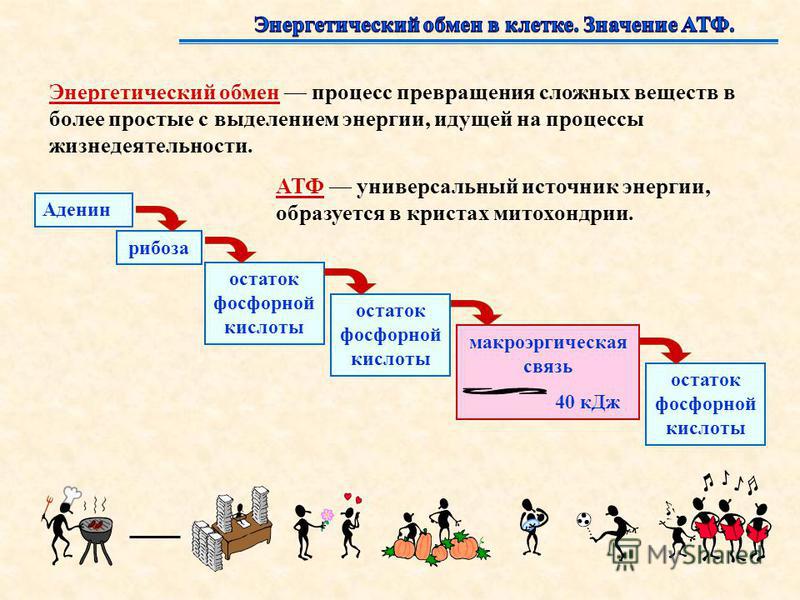 Конфумин разрешен к широкому медицинскому применению у взрослых, промышленный выпуск препарата освоен в ОАО «Фирма Медполимер».
Конфумин разрешен к широкому медицинскому применению у взрослых, промышленный выпуск препарата освоен в ОАО «Фирма Медполимер».
Energy Exchange — обзор
Анализ материальных и энергетических потоков
Материальные и энергетические обмены и сбережения начались в Калундборге в 1961 году, когда нефтеперерабатывающий завод Statoil начал использовать воду из озера Тиссо вместо грунтовых вод, экономя около 2 миллионов кубических метров воды в год . Затем Gyproc разместила свой завод в Калундборге, чтобы использовать топливный газ, поставляемый Statoil. К началу 1970-х годов нефтеперерабатывающий завод Statoil согласился поставлять свой избыточный газ (побочный продукт) компании Gyproc вместо того, чтобы сжигать его, что, по мнению Gyproc, является источником недорогого топлива.Позже, когда Statoil поставляла как очищенные сточные воды, так и охлаждающую воду на электростанцию Аснаес, она тем самым экономила в общей сложности 3 миллиона кубометров воды в год (вместо 2 миллионов), поскольку одна и та же вода «использовалась дважды» ».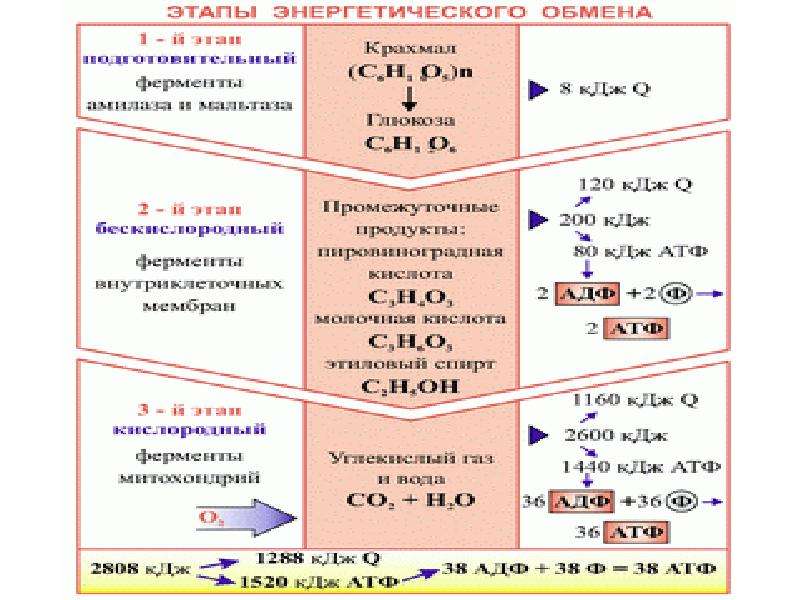 В 1976 году завод Ново Нордиск начал поток материалов, поставляя отстой от производственных процессов, а также от водоочистных сооружений рыбоводной фермы, который использовался в качестве удобрения для соседней фермы. Обмен шламов составил более 1 миллиона тонн в год.Кроме того, излишки дрожжей от производимого инсулина отправлялись фермерам в качестве корма для животных.
В 1976 году завод Ново Нордиск начал поток материалов, поставляя отстой от производственных процессов, а также от водоочистных сооружений рыбоводной фермы, который использовался в качестве удобрения для соседней фермы. Обмен шламов составил более 1 миллиона тонн в год.Кроме того, излишки дрожжей от производимого инсулина отправлялись фермерам в качестве корма для животных.
Производство ферментов основано на ферментации сырья, такого как картофельная мука и кукурузный крахмал. В процессе ферментации образуется около 150 000 кубических метров твердой биомассы, а также 90 000 кубических метров жидкой биомассы. За счет надлежащего перемещения этих отходов фермеры использовали их в качестве удобрений, тем самым сокращая потребление коммерческих удобрений.
Еще одна переработка отходов — это дрожжи, которые используются при производстве инсулина.Благодаря добавлению сахарной воды и молочной кислоты он превращается в корм для животных. Производство инсулина основано на процессе ферментации, в котором основными ингредиентами являются сахар и соль, которые превращаются в инсулин путем добавления дрожжей. После процесса нагревания дрожжи, остаточный продукт в этом производстве, превращаются в очень ценный корм: дрожжевую суспензию. К дрожжам добавляют сахарную воду и молочнокислые бактерии, что делает продукт более привлекательным для животных (800 000 свиней).
После процесса нагревания дрожжи, остаточный продукт в этом производстве, превращаются в очень ценный корм: дрожжевую суспензию. К дрожжам добавляют сахарную воду и молочнокислые бактерии, что делает продукт более привлекательным для животных (800 000 свиней).
Электростанция Аснаес работает на угле и работает с тепловым КПД около 40%, производя огромное количество энергии. Он использует соленую морскую воду для охлаждения, экономя воду озера Тиссо, и в то же время поставляет нагретую морскую воду в 57 близлежащих рыбоводных прудов, ежегодно производящих 200 тонн форели и лосося.
В 1981 году компания Asnaes начала снабжать районы паром для отопления, который заменил около 3 500 масляных печей и значительно снизил загрязнение воздуха.Кроме того, он снабжал паром компании Novo Nordisk и Statoil для процессов отопления. После того, как компания Statoil обработала избыточный газ путем удаления серы в соответствии с нормативными требованиями по выбросам серы, стало возможным использовать газ на электростанции Аснаес. Установка обессеривания Statoil снижает содержание серы в нефтеперерабатывающем газе, в результате чего выбросы SO 2 значительно сокращаются. Побочным продуктом является тиосульфат аммония, который используется в производстве примерно 20 000 тонн жидких удобрений, что примерно соответствует годовому потреблению в Дании.
Установка обессеривания Statoil снижает содержание серы в нефтеперерабатывающем газе, в результате чего выбросы SO 2 значительно сокращаются. Побочным продуктом является тиосульфат аммония, который используется в производстве примерно 20 000 тонн жидких удобрений, что примерно соответствует годовому потреблению в Дании.
В 1992 году электростанция Аснаес начала использовать очищенный газ компании Statoil вместо угля. Statoil также поставляет газ Gyproc в качестве источника энергии. Кроме того, удаленная сера продается в качестве сырья для производства серной кислоты на Камире. В 1993 году на электростанции Аснаес была добавлена установка обессеривания, которая удаляет серу из газов и производит сульфат кальция в виде отходов, известный как синтетический гипс. Обессеренная летучая зола используется цементной компанией, в то время как гипс поставляется Gyproc в качестве основного сырья для производства гипсокартона вместо импорта натурального гипса из Испании.В 1998 году электростанция производила приблизительно 190 000 тонн синтетического гипса в год.
Были также образованы и другие типы отходов, такие как 13 000 тонн газет / картона, которые после проверки качества продаются предприятиям, потребляющим картон и бумагу, в Дании, Швеции и Германии, производящим новую бумагу, новый картон, ящики для яиц и лотки. Еще 7000 тонн щебня и бетона были использованы для различных поверхностей после дробления и сортировки, и 15000 тонн садового / паркового мусора были доставлены в качестве мелиорации почвы в этом районе, а также 4000 тонн биологических отходов из домашних хозяйств и столовых компаний.Биоотходы используются в производстве компоста и биогаза. Четыре тысячи тонн железа и металла были перепроданы после очистки для вторичной переработки, а 1800 тонн стекла и бутылок были проданы производителям нового стекла.
С экологической точки зрения Калундборг демонстрирует характеристики простой пищевой сети: организмы потребляют отходы друг друга и энергию, тем самым становясь взаимозависимыми. Обмен повторно используемых и переработанных материалов и энергии из побочных продуктов промышленности привел к значительному увеличению прибыли и экономии средств. В течение 1993 года инвестиции в инфраструктуру (для транспортировки энергии и материалов) в размере 60 миллионов долларов принесли 120 миллионов долларов дохода и экономии средств. В 1998 году капитальные затраты на этот проект составили около 75 миллионов долларов. Экономия составила 160 миллионов долларов при сроке окупаемости менее 5 лет. При этом ежегодно экономятся десятки тысяч тонн воды, топлива и других продуктов. Снижение потребления природных ресурсов следующее: 45 000 тонн нефти / год, 15 000 тонн угля / год и 600 000 м 3 3 воды / год.Количество уменьшенных отходов и загрязнения также является значительным: 175 000 тонн углекислого газа в год; 10 200 тонн диоксида серы / год; 4500 тонн серы / год; 90 000 тонн сульфата кальция (гипса) / год; и 130 000 тонн летучей золы / год.
В течение 1993 года инвестиции в инфраструктуру (для транспортировки энергии и материалов) в размере 60 миллионов долларов принесли 120 миллионов долларов дохода и экономии средств. В 1998 году капитальные затраты на этот проект составили около 75 миллионов долларов. Экономия составила 160 миллионов долларов при сроке окупаемости менее 5 лет. При этом ежегодно экономятся десятки тысяч тонн воды, топлива и других продуктов. Снижение потребления природных ресурсов следующее: 45 000 тонн нефти / год, 15 000 тонн угля / год и 600 000 м 3 3 воды / год.Количество уменьшенных отходов и загрязнения также является значительным: 175 000 тонн углекислого газа в год; 10 200 тонн диоксида серы / год; 4500 тонн серы / год; 90 000 тонн сульфата кальция (гипса) / год; и 130 000 тонн летучей золы / год.
Хотя промышленный симбиоз Kalundborg был разработан благодаря деловому взаимодействию между компаниями, стремящимися экономно использовать свои побочные продукты и отходы, он принес как экономические, так и экологические преимущества. Обмен материалами осуществляется по замкнутому циклу, компании получают прибыль, а окружающая среда защищена за счет уменьшения загрязнения воздуха, воды и земли.
Обмен материалами осуществляется по замкнутому циклу, компании получают прибыль, а окружающая среда защищена за счет уменьшения загрязнения воздуха, воды и земли.
Тематическое исследование Kalundborg доказало, что устойчивость через промышленную экологию может быть прибыльной. Основным препятствием для внедрения методологии промышленной экологии является отсутствие лидерства в области промышленной экологии. Лидерство в области промышленной экологии является обязательным условием для разработки и поддержки методологии. Лидерство в области промышленной экологии должно осуществляться на общественных началах. Общение с высшим руководством производственной деятельности повысит коммуникативные навыки и разовьет взаимное доверие между партнерами, а также поможет людям общаться друг с другом и инициировать эко-промышленные системы.
Роль обмена поверхностной энергией для моделирования притока ветра: оценка нескольких моделей земной поверхности в WRF для отчета полевой кампании Южных Великих равнин (технический отчет)
Уортон, Соня, Симпсон, Мэтью, Осуна, Джессика, Ньюман, Дженнифер и Биро, Себастьян.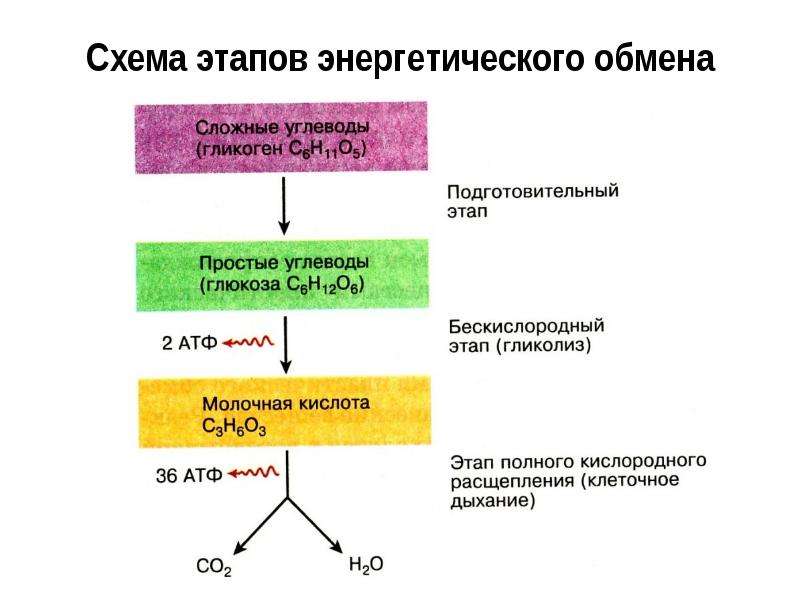 Роль обмена поверхностной энергией в моделировании притока ветра: оценка нескольких моделей земной поверхности в WRF для отчета полевой кампании Южных Великих равнин .США: Н. П., 2016.
Интернет. DOI: 10,2172 / 1254173.
Роль обмена поверхностной энергией в моделировании притока ветра: оценка нескольких моделей земной поверхности в WRF для отчета полевой кампании Южных Великих равнин .США: Н. П., 2016.
Интернет. DOI: 10,2172 / 1254173.
Уортон, Соня, Симпсон, Мэтью, Осуна, Джессика, Ньюман, Дженнифер и Биро, Себастьен. Роль обмена поверхностной энергией в моделировании притока ветра: оценка нескольких моделей земной поверхности в WRF для отчета полевой кампании Южных Великих равнин . Соединенные Штаты. https: // doi.org / 10.2172 / 1254173
Уортон, Соня, Симпсон, Мэтью, Осуна, Джессика, Ньюман, Дженнифер и Биро, Себастьян. Солнце .
«Роль обмена поверхностной энергией для моделирования притока ветра: оценка нескольких моделей земной поверхности в WRF для отчета полевой кампании Южных Великих равнин». Соединенные Штаты. https://doi.org/10.2172/1254173. https: //www.osti.гов / сервлеты / изнаночная / 1254173.
Соединенные Штаты. https://doi.org/10.2172/1254173. https: //www.osti.гов / сервлеты / изнаночная / 1254173.
@article {osti_1254173,
title = {Роль обмена поверхностной энергией в моделировании притока ветра: оценка нескольких моделей земной поверхности в WRF для отчета полевой кампании Южных Великих равнин},
author = {Уортон, Соня и Симпсон, Мэтью и Осуна, Джессика и Ньюман, Дженнифер и Биро, Себастьян},
abstractNote = {Модель исследования и прогнозирования погоды (WRF) используется для исследования выбора модели земной поверхности (LSM) для профиля приповерхностного ветра, включая высоты, достигаемые ветряными турбинами мощностью в несколько мегаватт.Моделирование профилей ветра и потоков поверхностной энергии было выполнено с использованием пяти LSM различной степени сложности для работы с обратными связями почва-растение-атмосфера для Департамента энергетики США (DOE) по измерению атмосферной радиации (ARM) Центра исследований климата Южных Великих равнин (SGP). ) Центральный объект в Оклахоме. Для проверки доступны измерения поверхностного потока и профиля ветра. Модель WRF использовалась в течение трех двухнедельных периодов, в течение которых существовали различные погодные условия и погодные условия.LSM предсказали широкий диапазон величин потоков энергии и сдвига ветра даже в прохладный осенний период, когда мы ожидали меньшей изменчивости. Моделирование потоков энергии различается по точности в зависимости от сложности модели, при этом LSM с очень простой обратной связью почва-растение-атмосфера или без нее были наименее точными; однако самые сложные модели не всегда давали более точные результаты. Ошибки в сдвиге ветра также были чувствительны к выбору LSM и частично были связаны с точностью данных о потоках энергии.Изменчивость характеристик LSM была относительно высокой, что свидетельствует о том, что представление LSM потоков энергии в модели WRF остается значительным источником неопределенности для моделирования условий притока ветровой турбины.
) Центральный объект в Оклахоме. Для проверки доступны измерения поверхностного потока и профиля ветра. Модель WRF использовалась в течение трех двухнедельных периодов, в течение которых существовали различные погодные условия и погодные условия.LSM предсказали широкий диапазон величин потоков энергии и сдвига ветра даже в прохладный осенний период, когда мы ожидали меньшей изменчивости. Моделирование потоков энергии различается по точности в зависимости от сложности модели, при этом LSM с очень простой обратной связью почва-растение-атмосфера или без нее были наименее точными; однако самые сложные модели не всегда давали более точные результаты. Ошибки в сдвиге ветра также были чувствительны к выбору LSM и частично были связаны с точностью данных о потоках энергии.Изменчивость характеристик LSM была относительно высокой, что свидетельствует о том, что представление LSM потоков энергии в модели WRF остается значительным источником неопределенности для моделирования условий притока ветровой турбины. },
},
doi = {10.2172 / 1254173},
url = {https://www.osti.gov/biblio/1254173},
журнал = {},
номер =,
объем =,
place = {United States},
год = {2016},
месяц = {5}
}
Энергетический обмен и экология по JSTOR
Окружающая среда имеет значение для организма через поток энергии и только через поток энергии.Энергия передается между окружающей средой и организмом посредством излучения, конвекции, теплопроводности и испарения влаги. Количественно описан обмен энергией между растением и окружающей средой. Дается описание климата растений, которое включает в себя радиацию, температуру воздуха, скорость ветра и относительную влажность. Важными свойствами растения являются впитывающая способность, размер листьев и сопротивление внутренней диффузии парообмену.
Изданный Американским институтом биологических наук (AIBS), BioScience представляет читателям своевременные и авторитетные обзоры текущих исследований в области биологии, сопровождаемые эссе и дискуссионными разделами по образованию, государственной политике, истории и концептуальным основам биологических наук. Рецензируемый, часто цитируемый ежемесячный журнал с содержанием, написанным и отредактированным для доступа исследователей, преподавателей и студентов, BioScience включает статьи о результатах и методах исследований, достижениях в области биологического образования, профессионально написанные тематические статьи о последних рубежах в биологии. , обсуждение профессиональных вопросов, обзоры книг, новости об AIBS, политическая колонка (Washington Watch) и образовательная колонка (Eye on Education). Круглые столы, форумы и статьи о точках зрения представляют точки зрения лидеров мнений и требуют дальнейших комментариев.BioScience входит в число лучших журналов в своей категории ISI (биология) как по импакт-фактору, так и по периодам полураспада цитирования.
Рецензируемый, часто цитируемый ежемесячный журнал с содержанием, написанным и отредактированным для доступа исследователей, преподавателей и студентов, BioScience включает статьи о результатах и методах исследований, достижениях в области биологического образования, профессионально написанные тематические статьи о последних рубежах в биологии. , обсуждение профессиональных вопросов, обзоры книг, новости об AIBS, политическая колонка (Washington Watch) и образовательная колонка (Eye on Education). Круглые столы, форумы и статьи о точках зрения представляют точки зрения лидеров мнений и требуют дальнейших комментариев.BioScience входит в число лучших журналов в своей категории ISI (биология) как по импакт-фактору, так и по периодам полураспада цитирования.
Oxford University Press — это отделение Оксфордского университета. Издание во всем мире способствует достижению цели университета в области исследований, стипендий и образования. OUP — крупнейшая в мире университетская пресса с самым широким присутствием в мире.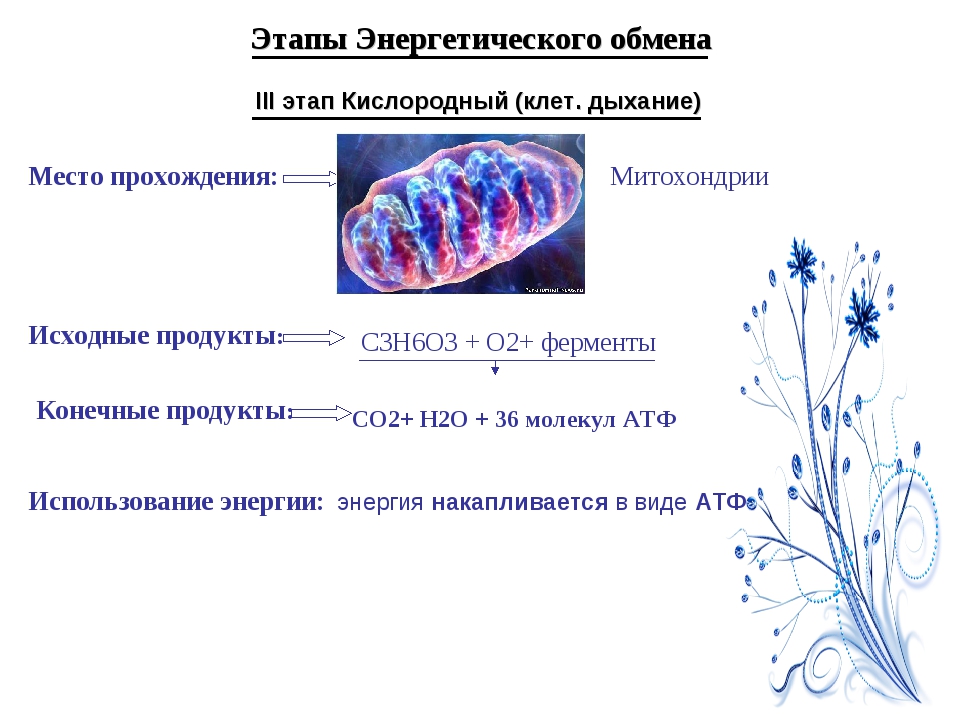 В настоящее время он издает более 6000 новых публикаций в год, имеет офисы примерно в пятидесяти странах и насчитывает более 5500 сотрудников по всему миру.Он стал известен миллионам людей благодаря разнообразной издательской программе, которая включает научные работы по всем академическим дисциплинам, библии, музыку, школьные и университетские учебники, книги по бизнесу, словари и справочники, а также академические журналы.
В настоящее время он издает более 6000 новых публикаций в год, имеет офисы примерно в пятидесяти странах и насчитывает более 5500 сотрудников по всему миру.Он стал известен миллионам людей благодаря разнообразной издательской программе, которая включает научные работы по всем академическим дисциплинам, библии, музыку, школьные и университетские учебники, книги по бизнесу, словари и справочники, а также академические журналы.
Условия использования веб-сайта HUDEX Energy Exchange Ltd.
Вы можете получить доступ к данным и информации (далее «Контент»), доступным на этом веб-сайте, только приняв эти условия использования; поэтому вас просят внимательно их прочитать.Если вы не принимаете эти условия, вы не сможете получить доступ к Контенту.
Любой контент, доступный на веб-сайте, предназначен исключительно для информационных целей; он не может и не должен рассматриваться как представляющий инвестиционный совет, исходящий от HUDEX Energy Exchange Ltd. , или публичное предложение в отношении любой из инвестиционных возможностей, представленных на веб-сайте.
, или публичное предложение в отношении любой из инвестиционных возможностей, представленных на веб-сайте.
HUDEX Energy Exchange Ltd. прилагает все усилия к тому, чтобы Контент, доступный на сервере, был правдивым, точным и своевременным, а доступ к веб-сайту был непрерывным и беспрепятственным, но HUDEX Energy Exchange Ltd.не гарантирует их и не несет ответственности за них. Пользователь несет исключительную ответственность за использование данных и информации.
Этот веб-сайт содержит ссылки на другие веб-сайты, обслуживаемые другими поставщиками услуг (ссылки). HUDEX Energy Exchange Ltd. не несет никакой ответственности за работу и содержание этих сайтов.
HUDEX Energy Exchange Ltd. не несет никакой ответственности за любые потери, убытки, затраты или расходы, возникшие, среди прочего, из-за ошибок, упущений или изменений в Содержании, доступном на сервере.
HUDEX Energy Exchange Ltd. рекомендует не принимать решения на основании транслируемого Контента до проверки информации. Пользователь несет полную ответственность за использование Контента. Как следствие, пользователь признает, что он / она использует Контент на свой страх и риск.
Пользователь несет полную ответственность за использование Контента. Как следствие, пользователь признает, что он / она использует Контент на свой страх и риск.
Содержимое этого веб-сайта предназначено для личного использования отдельными посетителями и может использоваться только в рамках бесплатного использования, предусмотренного законом. Любое дальнейшее распространение или распространение этого Контента, коммерческое или иное, любыми средствами массовой информации запрещено без письменного предварительного согласия HUDEX Energy Exchange Ltd.Воспроизведение, копирование, дублирование, передача или создание гипертекстовых ссылок любым способом всего или части Контента, который появляется на сайте, также требует предварительного письменного разрешения HUDEX Energy Exchange Ltd.
Если третья сторона, не являющаяся членом HUDEX, намеревается каким-либо образом включить Индекс Венгерских финансовых фьючерсов HUDEX в свои продукты или услуги, продаваемые на энергетической бирже или аналогичном предприятии, она должна заключить договор с HUDEX Energy Exchange Ltd. вышеуказанные данные (это положение не распространяется на внебиржевые операции).
вышеуказанные данные (это положение не распространяется на внебиржевые операции).
Несанкционированное или неправомерное использование или попытка несанкционированного или неправомерного использования Hungarian Financial Futures Index может повлечь за собой соответствующие судебные иски и уголовные или гражданские санкции. Передача Контента, обнаруженного на веб-сайте, неавторизованным третьим лицам считается несанкционированным и неправомочным использованием.
Все содержимое этого веб-сайта является собственностью HUDEX Energy Exchange Ltd., если не указано иное. Новости на этом веб-сайте можно использовать в рамках ограничений, предусмотренных законом, и их можно цитировать для предоставления информации.
Если вы используете через веб-сайт HUDEX Energy Exchange Ltd. любую услугу, где требуется ввод ваших личных данных, вы даете свое согласие на обработку таких данных. Персональные данные, предоставленные вами, будут использоваться HUDEX Energy Exchange Ltd. исключительно в тех целях, для которых вы их предоставили. HUDEX Energy Exchange Ltd. не будет передавать ваши личные данные третьим лицам.
исключительно в тех целях, для которых вы их предоставили. HUDEX Energy Exchange Ltd. не будет передавать ваши личные данные третьим лицам.
Большая межгодовая изменчивость CO 2 и энергообмена в пресноводных болотах при постоянных условиях окружающей среды
Резюме Мы проанализировали 5-летнюю запись CO 2 2 и энергообмена, надземной чистой первичной продукции (ANPP), максимального индекса площади листа ( LAI max ) и улучшенный индекс растительности (EVI) для болота Typha в Южной Калифорнии.На протяжении всего исследования болото было чистым источником углерода, несмотря на высокие уровни ANPP. Межгодовая изменчивость чистой продукции экосистемы (ЧЭП) была самой большой из всех, о которых сообщалось для любой наземной экосистемы, и объяснялась изменениями максимальной скорости фотосинтеза (GEE max ). Изменения в обмене энергии и массы были взаимосвязаны между годами; годы с уровнем поглощения углерода выше среднего были связаны с потоками явного тепла ниже среднего. Дистанционные измерения экологичности поверхности (EVI) были тесно связаны с вариациями GEE max , что является дополнительным свидетельством межгодовой изменчивости.Мы не смогли связать колебания GEE max с прямым воздействием погоды на физиологию экосистемы или межгодовой изменчивостью LAI max . GEE не изменялся систематически в зависимости от температуры воздуха или наличия стоячей воды на болоте; GEE max не отличался от LAI max в разные годы. Скорее, межгодовые колебания в обмене углерода на SJFM были вызваны изменениями в производственной эффективности болота (скорость валового или чистого обмена CO 2 на LAI), которые не были вызваны изменениями погоды.Наши результаты ставят под сомнение предположения о том, что межгодовые колебания в обмене сушей и атмосферой повсеместно вызваны прямым воздействием погоды на физиологию экосистемы и что физиологическая реакция экосистемы на физическую среду является постоянной из года в год.
Дистанционные измерения экологичности поверхности (EVI) были тесно связаны с вариациями GEE max , что является дополнительным свидетельством межгодовой изменчивости.Мы не смогли связать колебания GEE max с прямым воздействием погоды на физиологию экосистемы или межгодовой изменчивостью LAI max . GEE не изменялся систематически в зависимости от температуры воздуха или наличия стоячей воды на болоте; GEE max не отличался от LAI max в разные годы. Скорее, межгодовые колебания в обмене углерода на SJFM были вызваны изменениями в производственной эффективности болота (скорость валового или чистого обмена CO 2 на LAI), которые не были вызваны изменениями погоды.Наши результаты ставят под сомнение предположения о том, что межгодовые колебания в обмене сушей и атмосферой повсеместно вызваны прямым воздействием погоды на физиологию экосистемы и что физиологическая реакция экосистемы на физическую среду является постоянной из года в год.
Загрузить PDF для просмотраПросмотреть больше
Дополнительная информация Меньше информации
Закрывать
Введите пароль, чтобы открыть этот PDF-файл:
Отмена Ok
Подготовка документа к печати…
Отмена
(PDF) Обмен механической энергией во время ходьбы на самолете и преодоления препятствий у субъектов с синдромом Дауна
Ригольди, К. , Галли, М., и Альбертини, Г. (2010). Развитие походки в течение жизни у субъектов
, Галли, М., и Альбертини, Г. (2010). Развитие походки в течение жизни у субъектов
с синдромом Дауна. Исследование нарушений развития, 32: 158-63.
Смит Б. и Ульрих Б. (2008). Раннее применение стабилизирующих стратегий походки и препятствий:
пожилых человека с синдромом Дауна. Походка и осанка, 28 (3): 448-55.
Сазерленд Д. (2005). Эволюция клинического анализа походки, часть III — оценка кинетики и энергии
. Походка и поза, 21: 447-61.
Тейшейра-Салмела, Л., Надо, С., Милот, М., Гравел, Д. и Рекьяо, Л. (2008). Влияние каденции
на выработку и поглощение энергии в суставах нижних конечностей во время ходьбы
. Clinical Biomech, 23: 769-78.
Tesio, L., Roi, G., & Moller, F. (1991). Патологические походки: неэффективность — не правило. Clin
Biomech, 6: 47-50.
Ван де Валле, П., Халлеманс, А., Шварц, М., Труйен, С., Госселинк, Р., и Десловер, К.
(2011).Оценка механической энергии при ходьбе: достоверность и чувствительность при типичной походке
и у детей с церебральным параличом.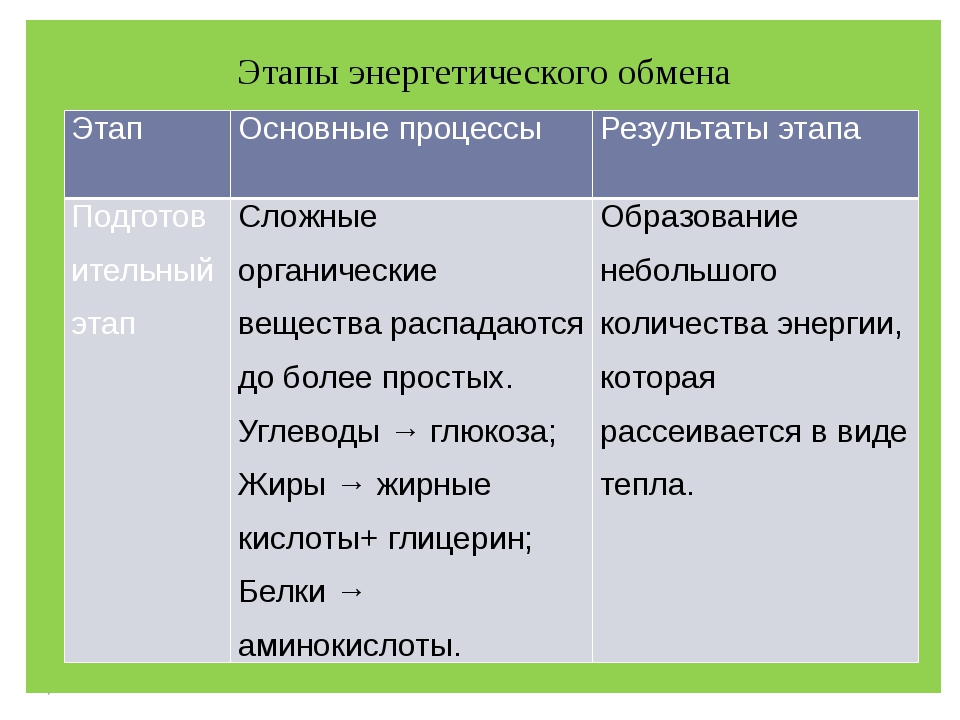 Походка и поза, 35 (2): 231-7.
Походка и поза, 35 (2): 231-7.
Вимеркати, С. Л., Галли, М., Ригольди, К., Анчиллао, А., и Альбертини, Г. (2011). Пространственно-временные
и кинетические аспекты уклонения от препятствий у субъектов с синдромом Дауна. J Mot
Behav.
Вимеркати, С., Галли, М., Ригольди, К., и Альбертини, Г. (2013). Избегание препятствий при синдроме Дауна
.Журнал электромиографии и кинезиологии, 23 (2): 483-9.
Вирджи-Бабул, Н., и Браун, М. (2004). Переход через препятствие: упреждающие модификации у
детей с синдромом Дауна и без него. Exp Brain Res, 159 (4): 487-90.
Уотерс, Р. Л., и Малрой, С. (1999). Энергозатраты нормальной и патологической походки.
Походка и поза, 9 (3): 207-31.
Weerdesteyn, V., Nienhuis, B., & Duysens, J. (2005). С возрастом возрастает
навыков избегания препятствий у пожилых людей.Hum Mov Sci., 24 (5-6): 865-80.
Изменение энергообмена между поверхностью и атмосферой и способность замораживания нижней зоны накопления, Западная Гренландия
Исследовательская статья 18 ноя 2015
Исследовательская статья | 18 ноя 2015
К. Харалампидис 1,2 , Д. фургон As 1 , J. E. Box 1 , M. Р. ван ден Брук 3 , W.Колган Т. 1,4 , С. Х. Дойл 5 , А. Л. Хаббард 6 , М. Макферрин 7 , H. Махгут 1,8 и К. Дж. П. Смитс 3 C. Charalampidis et al.
К. Харалампидис 1,2 , Д. фургон As 1 , J. E. Box 1 , M. Р. ван ден Брук 3 , W. Колган Т. 1,4 , С. Х. Дойл 5 , А. Л. Хаббард 6 , М. Макферрин 7 , H. Махгут 1,8 и К.Дж. П. П. Смитс 3
Харалампидис 1,2 , Д. фургон As 1 , J. E. Box 1 , M. Р. ван ден Брук 3 , W.Колган Т. 1,4 , С. Х. Дойл 5 , А. Л. Хаббард 6 , М. Макферрин 7 , H. Махгут 1,8 и К. Дж. П. Смитс 3 C. Charalampidis et al.
К. Харалампидис 1,2 , Д. фургон As 1 , J. E. Box 1 , M. Р. ван ден Брук 3 , W. Колган Т. 1,4 , С. Х. Дойл 5 , А. Л. Хаббард 6 , М. Макферрин 7 , H. Махгут 1,8 и К.Дж. П. П. Смитс 3 - 1 Геологическая служба Дании и Гренландии (GEUS), Øster Voldgade 10, 1350 Copenhagen K, Дания
- 2 Департамент наук о Земле, Университет Упсалы, Виллавэген 16, 752 36 Упсала, Швеция
- 3 Институт морских и атмосферных исследований (IMAU), Утрехтский университет, ПО Box 80005, 3508TA Утрехт, Нидерланды
- 4 Департамент наук о Земле и космосе, Йоркский университет, 4700 Keele Street, M3J 1P3, Торонто, Канада
- 5 Центр гляциологии Департамента географии и наук о Земле , Университет Аберистуита, Аберистуит, SY23 3DB, Великобритания
- 6 Центр арктических газовых гидратов, окружающей среды и климата, Департамент геологии, Университет Тромсё, Dramsveien 201, 9037 Тромсё, Норвегия
- 7 Кооперативный научно-исследовательский институт в Науки об окружающей среде (CIRES), 216 UCB, Университет Колорадо в Боулдере, Боулдер, Колорадо 80309, США
- 8 Центр арктических технологий (ARTEK), Технический университет Дании, Бровей, byg. 118, 2800 кг. Люнгбю, Дания
 118, 2800 кг. Люнгбю, Дания
118, 2800 кг. Люнгбю, Дания- 1 Геологическая служба Дании и Гренландии (GEUS), Øster Voldgade 10, 1350 Copenhagen K, Дания
- 2 Департамент наук о Земле, Университет Упсалы, Виллавэген 16, 752 36 Упсала, Швеция
- 3 Институт морских и атмосферных исследований (IMAU), Утрехтский университет, ПО Box 80005, 3508TA Утрехт, Нидерланды
- 4 Департамент наук о Земле и космосе, Йоркский университет, 4700 Keele Street, M3J 1P3, Торонто, Канада
- 5 Центр гляциологии Департамента географии и наук о Земле , Университет Аберистуита, Аберистуит, SY23 3DB, Великобритания
- 6 Центр арктических газовых гидратов, окружающей среды и климата, Департамент геологии, Университет Тромсё, Dramsveien 201, 9037 Тромсё, Норвегия
- 7 Кооперативный научно-исследовательский институт в Науки об окружающей среде (CIRES), 216 UCB, Университет Колорадо, Боулдер, Боулдер, CO 80309, США
- 8 Центр арктических технологий (ARTEK), Технический университет Дании, Бровей, byg. 118, 2800 кг. Люнгби, Дания
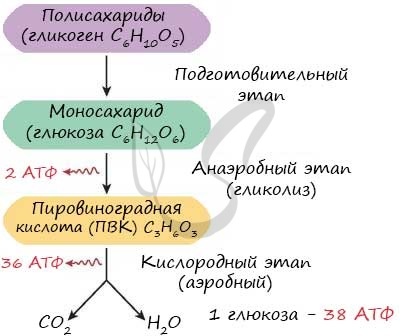 118, 2800 кг. Люнгби, Дания
118, 2800 кг. Люнгби, ДанияДля корреспонденции : К. Харалампидис ([email protected])
Скрыть данные об авторе Получено: 19 марта 2015 г. — Начало обсуждения: 27 мая 2015 г. — Исправлено: 18 октября 2015 г. — Принято: 25 октября 2015 г. — Опубликовано: 18 ноября 2015 г. Мы представляем 5 лет (2009–2013) автоматических измерений метеостанцией из нижней области накопления (1840 м над уровнем моря).- над уровнем моря) ледникового покрова Гренландии в районе Кангерлуссуак. Здесь оба лета 2010 и 2012 годов были исключительно теплыми, но только 2012 год привел к резко отрицательному балансу поверхностной массы (SMB) и поверхностному стоку талых вод. Наблюдаемый сток был связан с большой фракцией льда в верхних 10 м фирна, которая препятствовала просачиванию талой воды в доступный объем пор ниже. Анализ показывает аномально низкое среднее за лето 2012 г. альбедо 0,71 (обычно ~ 0,78), поскольку талая вода присутствовала на поверхности ледяного покрова. Следовательно, во время сезона таяния 2012 года поверхность ледникового покрова поглотила на 28% (213 МДж м −2 ) больше солнечной радиации, чем в среднем за все другие годы.
Следовательно, во время сезона таяния 2012 года поверхность ледникового покрова поглотила на 28% (213 МДж м −2 ) больше солнечной радиации, чем в среднем за все другие годы.
Модель баланса поверхностной энергии используется для оценки сезонной и межгодовой изменчивости всех потоков поверхностной энергии. Модель воспроизводит наблюдаемые скорости таяния, а также SMB для каждого сезона. Анализ чувствительности показывает, что 71% дополнительной солнечной радиации в 2012 году было использовано для плавления, что соответствует 36% (0.64 м) спуска 2012 г. Остальные 64% (1,14 м) понижения поверхности были вызваны высокими температурами атмосферы, до +2,6 ° C в среднем за сутки, что указывает на то, что 2012 год был бы отрицательным годом SMB на этом участке даже без обратной связи таяние – альбедо.
Более длинные временные ряды SMB, региональной температуры и альбедо дистанционного зондирования (MODIS) показывают, что 2012 год был первым годом сильно отрицательного SMB с самым низким альбедо на этой высоте за всю историю наблюдений.