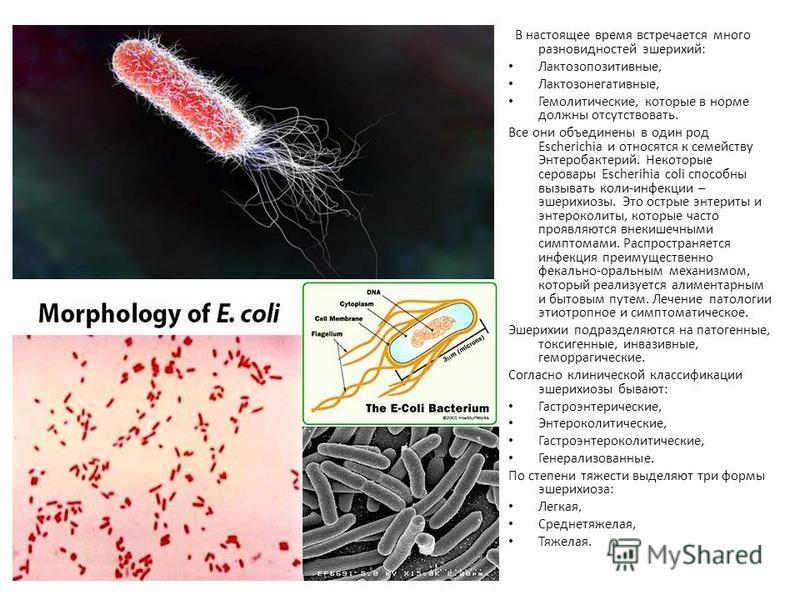
Кишечная палочка (E.coli)
СТАЙЛАБ предлагает тест-системы для определения кишечной палочки (E. coli) в пробах продуктов и окружающей среды микробиологическими методами, с помощью ПЦР в реальном времени, а также методом латекс-агглютинации.
Кишечная палочка (Escherichia coli) – это факультативно-анаэробная грамотрицательная подвижная палочковидная бактерия, входящая в состав нормальной микрофлоры кишечника большинства теплокровных животных, в том числе, и человека. Существует множество серотипов кишечной палочки, большинство из которых безвредно или даже полезно для носителя. Бактерии, входящие в состав микрофлоры, препятствуют размножению других бактерий, в том числе, патогенных. Кроме того, кишечная палочка вырабатывает витамин К. Однако некоторые серотипы E. coli могут вызывать тяжелые заболевания. Морфологически безвредные и патогенные кишечные палочки не различаются, потому для определения патогенности микроорганизма необходимо проанализировать его антигены.
Бактерии кишечной палочки могут относиться к различным серогруппам: обладать различными антигенами. Серогруппа – это группа бактерий, обладающих общим антигеном; она может включать более одного серотипа, вида или рода. Для E. coli наиболее известными являются серогруппы энтеропатогенной кишечной палочки O26, O55, O86, O111, O114, O119, O125, O126, O127, O128, O142 и O158. К этим же группам могут относиться и другие типы патогенной кишечной палочки. Например, у DAEC выявлены серогруппы O86, O127, O142 и O158.
Выделяют шесть типов патогенных кишечных палочек.
- Энтерогеморрагическая кишечная палочка (EHEC, используются также сокращения STEC – от «шигатоксин-продуцирующая E. coli и VTEC – «веротоксин-продуцирующая E.Coli). Эти бактерии вырабатывают два токсина, которые называют веротоксинами, или шигаподобными (шигеллоподобными) токсинами. EHEC вызывает заболевания, по симптоматике сходные с дизентерией и сопровождающиеся геморрагической диареей.
 В тяжелых случаях может развиваться гемолитико-уремический синдром (ГУС). К EHEC относятся серотипы кишечной палочки O104:h5, O157:H7 и некоторые другие.
В тяжелых случаях может развиваться гемолитико-уремический синдром (ГУС). К EHEC относятся серотипы кишечной палочки O104:h5, O157:H7 и некоторые другие. - Энтеротоксигенная кишечная палочка (ETEC) вырабатывает термолабильный и термостабильный токсины, последний из которых похож на токсин холерного вибриона. Для заболеваний, вызываемых этой бактерией, характерна водянистая диарея, повышенная температура и, в некоторых случаях, тошнота.
- Энтероинвазивная кишечная палочка (EIEC) вызывает заболевания, похожие на бактериальную дизентерию. Эти бактерии проникают в клетки эпителия кишечника и размножаются в них.
- Энтеропатогенная кишечная палочка (EPEC) чаще вызывает диарею у детей, чем у взрослых. Механизм ее действия отличается от ETEC и EIEC: эти бактерии прикрепляются к клеткам эпителия кишечника, но не проникают в них. Заболевания, вызванные EPEC, могут продолжаться до 2 недель.
- Энтероагрегативная кишечная палочка (EAEC) также вызывает заболевания, в основном, у детей. Бактерии прикрепляются к эпителию кишечника и выделяют токсины.
- Диффузно-адгезивная кишечная палочка (DAEC) вызывает легкую диарею у детей, особенно возрастом до 2х лет. Однако этот микроорганизм обнаруживали и у здоровых детей, а также у здоровых взрослых.
 В тяжелых случаях может развиваться гемолитико-уремический синдром (ГУС). К EHEC относятся серотипы кишечной палочки O104:h5, O157:H7 и некоторые другие.
В тяжелых случаях может развиваться гемолитико-уремический синдром (ГУС). К EHEC относятся серотипы кишечной палочки O104:h5, O157:H7 и некоторые другие. Бактерии прикрепляются к эпителию кишечника и выделяют токсины.
Бактерии прикрепляются к эпителию кишечника и выделяют токсины.Кишечная палочка способна вызывать не только энтериты и токсикоинфекции, но и менингиты новорожденных, респираторные инфекции, инфекции мочевыводящих путей и бактериемию – общее поражение организма, вызванное бактериями, проникшими в кровь.
E. coli относится к наиболее изученным организмам. Геном одного из штаммов этих бактерий был полностью секвенирован в 1997 году. Кишечную палочку используют в генетических и микробиологических экспериментах, а также при изучении видообразования. Кроме того, с помощью этих бактерий синтезируют различные белки, в частности, аналог инсулина человека, некоторые ферменты и др. Их применяют также для получения вакцин. Ведутся исследования по использованию кишечной палочки для получения топлива.
В Российской Федерации и странах Таможенного Союза содержание кишечной палочки в пищевых продуктах ограничено ТР ТС 021/2011 «О безопасности пищевой продукции», ТР ТС 033/2013 «О безопасности молока и молочной продукции» и другими техническими регламентами. С актуальной законодательной информацией можно ознакомиться на сайте
Для определения кишечной палочки в пробах используют как микробиологические методы (селективные среды, тест-подложки), так и определение ДНК с помощью ПЦР. В качестве экспресс-тестов применяют простой и быстрый метод латекс-агглютинации.
Литература
- О.К. Поздеев. Медицинская микробиология. Москва, ГЭОТАР-МЕД, 2001.
- Официальный сайт CDC.
- Germani Y, Bégaud E, Duval P, Le Bouguénec C. Prevalence of enteropathogenic, enteroaggregative, and diffusely adherent Escherichia coli among isolates from children with diarrhea in new Caledonia. J Infect Dis. 1996 Nov;174(5):1124-6.
- Mansan-Almeida R, Pereira AL, Giugliano LG. Diffusely adherent Escherichia coli strains isolated from children and adults constitute two different populations. BMC Microbiol. 2013 Feb 1;13:22.
- Isabel C. A. Scaletsky, Sandra H. Fabbricotti, Rozane L. B. Carvalho, Claudia R. Nunes, Helcio S. Maranhão, Mauro B. Morais, and Ulysses Fagundes-Neto Diffusely Adherent Escherichia coli as a Cause of Acute Diarrhea in Young Children in Northeast Brazil: a Case-Control Study. J Clin Microbiol. 2002 Feb; 40(2): 645–648.
 1996 Nov;174(5):1124-6.
1996 Nov;174(5):1124-6.общее микробное число
ХАРАКТЕРИСТИКА КЛИНИЧЕСКИХ ИЗОЛЯТОВ ESCHERICHIA COLI, ВЫДЕЛЕННЫХ ОТ ПАЦИЕНТОВ С БОЛЕЗНЬЮ КРОНА | Городничев
1. Ahmed I., Roy В. С., Khan S. A. et al. Microbiome, metabolome and inflammatory bowel disease. Microorganisms. 2016, 4 (2): 20.
2. Baumgart М., Dogan B., Rishniw M. et al. Culture independent analysis of ileal mucosa reveals a selective increase in invasive Escherichia coli of novel phylogeny relative to depletion of clostridiales in Crohn’s disease involving the ileum. ISME J. 2007, 1 (5): 403-418.
Baumgart М., Dogan B., Rishniw M. et al. Culture independent analysis of ileal mucosa reveals a selective increase in invasive Escherichia coli of novel phylogeny relative to depletion of clostridiales in Crohn’s disease involving the ileum. ISME J. 2007, 1 (5): 403-418.
3. Carriere J., Darfeuille-Michaud A., Nguyen H. T. Infectious etiopathogenesis of Crohn’s disease. World J. Gastroenterol. 2014, 20 (34): 12102-12117.
4. Comito D., Cascio A., Romano C. Microbiota biodiversity in inflammatory bowel disease. Italian J. Pediatrics. 2014, 40 (1): 1.
5. Clermont O., Bonacorsi S., Bingen E. Rapid and simple determination of the Escherichia coli phylogenetic group. Applied Environmental Microbiology. 2000, 66 (10): 4555-4558.
6. Liang X., Ji Y. Comparative analysis of staphylococcal adhesion and internalization by epithelial cells. Methicillin-resistant Staphylococcus aureus (MRSA) protocols. 2007, p. 145-151.
Liang X., Ji Y. Comparative analysis of staphylococcal adhesion and internalization by epithelial cells. Methicillin-resistant Staphylococcus aureus (MRSA) protocols. 2007, p. 145-151.
7. Martinez-Medina М., Garcia-Gil L.J. Escherichia coli in chronic inflammatory bowel diseases: An update on adherent invasive Escherichia coli pathogenicity. World J, Gastrointest. Pathophysiol. 2014, 5 (3): 213-227.
8. Martinez-Medina М., Aldeguer X., Lopez-Siles M. et al. Darfeuille-michaud a molecular diversity of Escherichia coli in the human gut: new ecological evidence supporting the role of adherent-invasive E. coli (AIEC) in Crohn’s disease. Inflammatory Bowel Diseases. 2009, 15 (6): 872-882.
9. Wood Т.К., Barrios A.F.G., Herzberg М., Lee J. Motility influences biofilm architecture in Escherichia coli.
Урология » Сравнительная характеристика штаммов уропатогенной Escherichia coli, выделенных в условиях поликлиники и стационара
1. Grabe M., Bjerklund-Johansen T.E., Botto H., Cek M., Naber K.G., Pickard R.S.,Tenke P., Wagenlehner F., Wullt B. Urological infections. European Guidelines. 2011;115 p.
2. Wiles T.J., Kulesus R.R., Mulvey M.A. Origins and virulence mechanisms of uropathogenic Escherichia coli. Exp. Мol. Рathol. 2008;5:11–19.Doi:10.1016/j.yexmp.2008.03.007.
3. Soto S.M. Importance of biofilms in urinary tract infections: new therapeutic approaches. Adv. Biol. 2014;13:543974. ID 543974.
4. Nicolle L.E. Catheter associated urinary tract infections. Antimicrob. Resist. Inf. Control. 2014;3(23):1–8. Doi: 10.1186/2047-2994-3-23.
5. Hola V., Ruzicka F. The Formation of poly-microbial biofilms on urinary catheters. urinary tract infections, Dr. Peter Tenke (Ed.), ISBN: 978-953-307-757-4, InTech Available from: http://www.intechopen.com/books/urinary-tract-infections/the-formation-of-poly-microbial-biofilms-on-urinarycatheters.
Peter Tenke (Ed.), ISBN: 978-953-307-757-4, InTech Available from: http://www.intechopen.com/books/urinary-tract-infections/the-formation-of-poly-microbial-biofilms-on-urinarycatheters.
6. Bukharin O.V., Gritsenko V.A., Vyalkova A.A. Factors of uropathogenicity of bacteria: role in pathogenesis and importance in diagnosis of pyelonephritis. Nephrology and dialysis. 2001;3(4):469–475. Russian (Бухарин О.В., Гриценко В.А., Вялкова А.А. Факторы уропатогенности бактерий: роль в патогенезе и значение в диагностике пиелонефрита. Нефрология и диализ. 2001;3(4):469–475).
7. Perepanova T.S. The importance of infections caused by the formation of biofilms in urological practice. Effective pharmacotherapy. 2013;37:18–27. Russian (Перепанова Т.С. Значение инфекций, обусловленных образованием биопленок, в урологической практике. Эффективная фармакотерапия. 2013;37:18–27).
8. Kanamaru S., Kurazono H., Nakano M., Terai A., Ogawa O., Yamamoto S. Subtyping of uropathogenic Escherichia coli according to the pathogenicity island encoding uropathogenic-specific protein: comparison with phylogenetic groups. Int. J. Urol. 2006;13:754–760. Doi: 10.1111/j.1442-2042.2006.01398.x.
Int. J. Urol. 2006;13:754–760. Doi: 10.1111/j.1442-2042.2006.01398.x.
9. Lee J.H., Subhadra B., Kim D.H., Park H.S., Kim J.M., Koo S.H., Oh M.H.,Choi C.H. Phylogenetic group distributions, virulence factors and antimicrobial resistance properties of uropathogenic Escherichia coli strains isolated from patients with urinary tract infections in South Korea. Lett. App. Microbiol. 2015;62:84–90. Doi: 10.1111/lam.12517.
10. Millán Y., Hernández E., Millán B., Araque M. Distribution of phylogenetic groups and virulence factors in CTX-M-15 β-lactamase-producing uropathogenic Escherichia coli strains isolated from patients in the community of Mérida, Venezuela. Rev. Argent. Microbiol. 2014;46(3):175–81. Doi: 10.1016/S0325-7541(14)70069-0.
11. Munkhdelger Y. Detection of virulence genes, phylogenetic group and antibiotic resistance of uropathogenic Escherichia coli in Mongolia. J. Infect. Dev. Ctries. 2017;11(1):51–57. Doi: 10.3855/jidc.7903.
12. Grude N., Potaturkina-Nesterovа N. I., Jenkins A., Strand L., Nowrouzian F.L.,Nyhus J., Kristiansen B.E. A comparison of phylogenetic group, virulence factors and antibiotic resistance in Russian and Norwegian isolates of Escherichia coli from urinary tract infection. Clinical Microbiology and Infection. 2007;13(2):208–211. Doi: 10.1111/j.1469-0691.2006.01584.x.
I., Jenkins A., Strand L., Nowrouzian F.L.,Nyhus J., Kristiansen B.E. A comparison of phylogenetic group, virulence factors and antibiotic resistance in Russian and Norwegian isolates of Escherichia coli from urinary tract infection. Clinical Microbiology and Infection. 2007;13(2):208–211. Doi: 10.1111/j.1469-0691.2006.01584.x.
13. Kumar N., Nahid F., Zahra R. Association of virulence factors, phylogenetic groups and antimicrobial resistance markers in Escherichia coli from Badin city, Pakistan. J. Chemother. 2017;29(1):8–13. Doi:10.1080/1120009X.2016.1154682.
14. O’Toole G.A., Kolter R. Flagellar and twitching motility are necessary for Pseudomonas aeruginosa biofilm development. Mol. Microbiol. 1998;30:295–304. Doi: 10.1046/j.1365-2958.1998.01062.x.
15. Clermont O., Christenson J.K., Denamur E., Gordon D.M. The Clermont Escherichia coli phylo-typing method revisited: improvement of specificity and detection of new phylo-groups. Environ. Microbiol. Rep. 2013;1:58–61. Doi: 10.1111/1758-2229.12019.
Doi: 10.1111/1758-2229.12019.
16. Flores-Mireles A.L., Walker J.N., Caparon M., Hultgren S.J. Urinary tract infections: epidemiology, mechanisms of infection and treatment options. Nat. Rev. Microbiol. 2015; 13(5):269–284. Doi:10.1038/nrmicro3432.
17. Katouli M. Population structure of gut Escherichia coli and its role in development of extra-intestinal infections. Iran. J. Microbiol. 2010;2(2):59–72.
18. Kogan M.I., NabokaYu.L., Bedzhanyan S.K., Mitusova E.V., Gudima I.A., Morgun P.P., Vasil’eva L.I. Is bacteriological testing of bladder urine informative in acute obstructive pyelonephritis? Urologiia. 2017;3:10–15. Russian (Коган М.И., Набока Ю.Л., Беджанян С.К., Митусова Е.В.,Гудима И.А., Моргун П.П., Васильева Л.И. Информативно ли бактериологическое исследование пузырной мочи при остром обструктивном пиелонефрите? Урология. 2017;3:10–15). Doi: https://dx.Doi.org/10.18565/urol.2017.3.10-15.
19. Shamkhalova M.Sh., Chugunova L.A. Urinary tract infections in diabetic patients: diagnosis, prevention, treatment. Guidelines. International Endocrinology Journal. 2005; 2(2)
Guidelines. International Endocrinology Journal. 2005; 2(2)
20. Kline K.A., Lewis A.L. Gram-positive uropathogens, polymicrobial urinary tract infection, and the emerging microbiota of the urinary tract. Microbiol. Spectr. 2016;4(2). Doi: 10.1128/microbiolspec.UTI-0012-2012.
21. Matthews S.J., Lancaster J.W. Urinary tract infections in the elderly population. Am. J. Geriatr. Pharmacother. 2011;9:286–309.
22. Warren J.W., Tenney J.H., Hoopes J.M., Muncie H.L., Anthony W.C.А prospective microbiologic study of bacteriuria in patients with chronic indwelling urethral catheters. J. Infect. Dis. 1982;146(6):719–723. Doi: 10.1093/infdis/146.6.719.
23. Galvan E.M., Mateyca C., Ielpi L. Role of interspecies interactions in dual-species biofilms developed in vitro by uropathogens isolated from polymicrobial urinary catheter associated bacteriuria. Biofouling. 2016;32(9):1067–1077. Doi: 10.1080/08927014.2016.1231300.
24. Bishara J., Leibovici L., Huminer D. , Drucker M., Samra Z., Konisberger H., Pitlik S. Five-year prospective study of bacteraemic urinary tract infection in a single institution. Eur. J Clin. Microbiol. Infect. Dis. 1997;16(8):563–567.
, Drucker M., Samra Z., Konisberger H., Pitlik S. Five-year prospective study of bacteraemic urinary tract infection in a single institution. Eur. J Clin. Microbiol. Infect. Dis. 1997;16(8):563–567.
25. Tets G.V., Tets V.V., Voroshilova T.M. Metagenomic analysis of samples in cystitis. Urologiia. 2016; 1:37–44. Russian (Тец Г.В., Тец В.В., Ворошилова Т.М. Метагеномный анализ проб при цистите. Урология. 2016; 1:37–44).
26. Tenaillon O., Skurnik D., Picard B., Denamur E. The population genetics of commensal Escherichia coli. Nat. Rev. Microbiol. 2010;8(3):207–217. Doi:10.1038/nrmicro2298.
27. Clermont O., Bonacorsi S., Bingen E. Rapid and simple determination of the Escherichia coli phylogenetic group. Appl. Environ. Microbiol. 2000;10:4555–4558. Doi: 10.1128/AEM.66.10.4555-4558.2000.
28. Logue C.M., Wannemuehler Y., Nicholson B.A., Doetkott C., Barbieri N.L., Nolan L.K. Comparative analysis of phylogenetic assignment of human and avian ExPEC and fecal commensal Еscherichia coli using the (previous and revised) Clermont phylogenetic typing methods and its impact on avian pathogenic Escherichia coli (APEC) classification. Front. Microbiol. 2017;8:283. Doi:10.3389/fmicb.2017.00283.
Front. Microbiol. 2017;8:283. Doi:10.3389/fmicb.2017.00283.
29. Iranpour D., Hassanpour M., Ansari H., Tajbakhsh S., Khamisipour G., Najafi A. Phylogenetic groups of Escherichia coli strains from patients with urinary tract infection in Iran based on the new Clermont phylotyping method. Bioмed. Res. Int. 2015;846219. ID 846219. Doi: 10.1155/2015/846219.
30. Skjøt-Rasmussen L., Ejrnæs K., Lundgren B., Frimodt-Møller N. Virulence factors and phylogenetic grouping of Escherichia coli isolates from patients with bacteraemia of urinary tract origin relate to sex and hospital- vs. community-acquired origin. Int. J. Med. Microbiol. 2012;302(3):129–134. Doi: 10.1016/j.ijmm.2012.03.002.
31. Turrientes M.-C., González-Alba J.-M., del Campo R., Baquero M.-R., Cantón R., Baquero F., Galan J.-C. Recombination blurs phylogenetic groups routine assignment in Escherichia coli: setting the record straight. PLoS One. 2014;9(11):e113532.
32. Fatima N., Agrawal M., Shukla I. , Khan P.A. Characterization of uropathogenic E. coli in relation to virulence factors. Scientific Reports. 2012;1(7):342. Doi:10.4172/scientificreports.
, Khan P.A. Characterization of uropathogenic E. coli in relation to virulence factors. Scientific Reports. 2012;1(7):342. Doi:10.4172/scientificreports.
33. Ejrnæs K., Stegger M., Reisner A., Ferry S., Monsen T., Holm S.E., Lundgren B.,Frimodt-Møller N. Characteristics of Escherichia coli causing persistence or relapse of urinary tract infections: phylogenetic groups, virulence factors and biofilm formation. Virulence. 2011;2(6):528–537. Doi: 10.4161/viru.2.6.18189.
34. Palagin I.S., Sukhorukova M.V., Dekhnich A.V., Eydelshteyn M.V., Shevelev A.N. et al. The current state of antibiotic resistance of pathogens of community-acquired urinary tract infections in Russia: results of the study «DARMIS» (2010–2011). Klin. Microbiol. Antimicrobial. Khimioter. 2012;14(4):280–302. Russian (Палагин И.С., Сухорукова М.В., Дехнич А.В.,Эйдельштейн М.В., Шевелев А.Н, Гринев А.В., Перепанова Т.С., Козлов Р.С. Современное состояние антибиотикорезистентности возбудителей внебольничных инфекций мочевых путей в России: результаты исследования «ДАРМИС» (2010–2011). Клиническая микробиология и антимикробная химиотерапия. 2012;14(4):280–302).
Клиническая микробиология и антимикробная химиотерапия. 2012;14(4):280–302).
35. Ali I., Kumar N., Ahmed S., Dasti J.I. Antibiotic resistance in uropathogenic E. сoli strains isolated from non-hospitalized patients in Pakistan. J. Clin. Diagn. Res. 2014;8(9):1–4. Doi:10.7860/JCDR/2014/7881.4813.
36. Rijavec M., Erjavec Starčič M., Avguštin Ambrožič J., Reissbrodt R., Fruth A., Križan-Hergouth V., Žgur-Bertok D. High prevalence of multidrug resistance and random distribution of mobile genetic elements among uropathogenic Escherichia coli (UPEC) of the four major phylogenetic groups. Curr. Microbiol. 2006;53:158–162. Doi:10.1007/s00284-005-0501-4.
37. Petty N.K., Ben Zakour N.L., Stanton-Cook M., Skippington E., Totsika M., Forde B.M., Phan M.D. et al. Global dissemination of a multidrug resistant Escherichia coli clone. Proc. Natl. Acad. Sci. USA. 2014;111(15):5694–5699. Doi:10.1073/pnas.1322678111.
Протеобактерии — Proteobacteria — qaz.wiki
Тип грамотрицательных бактерий
Proteobacteria является одним из основных фила из грам-отрицательных бактерий . Они включают в себя широкий спектр патогенных родов, таких как Escherichia , Salmonella , Vibrio , Helicobacter , Yersinia , Legionellales и многие другие. Другие являются свободноживущими (не паразитическими ) и включают многие бактерии, ответственные за фиксацию азота .
Они включают в себя широкий спектр патогенных родов, таких как Escherichia , Salmonella , Vibrio , Helicobacter , Yersinia , Legionellales и многие другие. Другие являются свободноживущими (не паразитическими ) и включают многие бактерии, ответственные за фиксацию азота .
Карл Вёзе основал эту группировку в 1987 году, неофициально назвав ее «пурпурные бактерии и их родственники». Из-за большого разнообразия форм, встречающихся в этой группе, он был назван в честь Протея , греческого бога моря, способного принимать самые разные формы, и не назван в честь рода Proteus .
Характеристики
Все «Протеобактерии» являются грамотрицательными (хотя некоторые из них могут окрашивать грамположительные или грамположительные на практике), их внешняя мембрана в основном состоит из липополисахаридов . Многие передвигаются, используя жгутики , но некоторые неподвижны или полагаются на скольжение бактерий . К последним относятся миксобактерии — отряд бактерий, которые могут объединяться с образованием многоклеточных плодовых тел. Также существует большое разнообразие типов метаболизма . Большинство членов являются факультативно или обязательно анаэробными , хемолитоавтотрофными и гетеротрофными , но встречаются многочисленные исключения. Множество родов, которые не связаны между собой близко друг к другу, преобразуют энергию света посредством фотосинтеза и аноксигенного фотосинтеза .
К последним относятся миксобактерии — отряд бактерий, которые могут объединяться с образованием многоклеточных плодовых тел. Также существует большое разнообразие типов метаболизма . Большинство членов являются факультативно или обязательно анаэробными , хемолитоавтотрофными и гетеротрофными , но встречаются многочисленные исключения. Множество родов, которые не связаны между собой близко друг к другу, преобразуют энергию света посредством фотосинтеза и аноксигенного фотосинтеза .
«Протеобактерии» связаны с дисбалансом микробиоты нижних половых путей женщины. Эти виды связаны с воспалением.
Некоторые Alphaproteobacteria могут расти при очень низком уровне питательных веществ и иметь необычную морфологию, например стебли и почки. Другие включают важные в сельском хозяйстве бактерии, способные вызывать азотфиксацию в симбиозе с растениями. Порядок типов — Caulobacterales , включающий стеблеобразующие бактерии, такие как Caulobacter . В митохондриях эукариот, как считается потомками alphaproteobacterium.
В митохондриях эукариот, как считается потомками alphaproteobacterium.
Betaproteobacteria весьма разнообразен метаболический и содержит chemolithoautotrophs , фотоавтотроф и общие профиль гетеротроф . Порядок типов — Burkholderiales , включающий огромное разнообразие метаболических микроорганизмов , включая условно-патогенные микроорганизмы .
Гамма-протеобактерии являются самым крупным классом с точки зрения видов, правомерно опубликованных названий. Порядок типов — Pseudomonadales , который включает роды Pseudomonas и азотфиксирующие Azotobacter .
Deltaproteobacteria включают бактерии , которые являются хищниками на другие бактерии и являются важными факторами , способствующими анаэробной части серного цикла. Порядок типов — Myxococcales , который включает организмы со способностями к самоорганизации, такие как Myxococcus spp.
Epsilonproteobacteria часто являются тонким, грамотрицательными палочками , которые являются спиральными или изогнутыми. Порядок типов — Campylobacterales , который включает важные пищевые патогены, такие как Campylobacter spp.
Порядок типов — Campylobacterales , который включает важные пищевые патогены, такие как Campylobacter spp.
Zetaproteobacteria являются железо-окисляющих нейтрофильные chemolithoautotrophs , распределенных по всему миру в эстуарии и морской среды обитания. Типовой заказ — Марипрофундалес .
Hydrogenophilalia являются облигатными термофилами и включают в себя гетеротроф и автотроф. Порядок типа — Hydrogenophilales .
Acidithiobacillia содержат только серу, железо и уран-окислительных автотроф . Порядок типов — Acidithiobacillales , который включает экономически важные организмы, используемые в горнодобывающей промышленности, такие как Acidithiobacillus spp.
Oligoflexia являются нитевидными аэробами. Порядок типов — Oligoflexales , который содержит род Oligoflexus .
Таксономия
| Филогения «протеобактерий». |
Филогения «Proteobacteria» по данным ARB living tree, iTOL, Bergey’s и др. Ацидобактерии (не путать с ацидитиобациллами) используются в качестве внешней группы. Ацидобактерии (не путать с ацидитиобациллами) используются в качестве внешней группы. |
Группа определяется в первую очередь с точки зрения последовательностей рибосомной РНК (рРНК). «Протеобактерии» разделены на девять классов с официально опубликованными названиями, обозначаемыми греческими буквами от альфа до дзета, Acidithiobacillia, Hydrogenophilalia и Oligoflexia. Раньше они считались подклассами этого типа, но теперь рассматриваются как классы . Эти классы монофилетичны . Род Acidithiobacillus , входивший в состав Gammaproteobacteria до тех пор, пока в 2013 году он не был переведен в класс Acidithiobacillia, ранее считался парафилетическим по отношению к бетапротеобактериям согласно исследованиям мультигеномного выравнивания . В 2017 году Betaproteobacteria подверглись серьезным изменениям, и был создан класс Hydrogenophilalia, содержащий отряд Hydrogenophilales.
Классы протеобактерий с достоверно опубликованными названиями включают некоторые известные роды: например:
- Альфапротеобактерии : Brucella , Rhizobium , Agrobacterium , Caulobacter , Rickettsia , Wolbachia и др.
- Бетапротеобактерии : Bordetella , Ralstonia , Neisseria , Nitrosomonas и др.
- Гаммапротеобактерии : Escherichia , Shigella , Salmonella , Yersinia , Buchnera , Haemophilus , Vibrio , Pseudomonas и др.
- Дельтапротеобактерии : Desulfovibrio , Geobacter , Bdellovibrio и др.
- Эпсилонпротеобактерии : Helicobacter , Campylobacter , Wolinella и др.
- Зетапротеобактерии : Марипрофундус , Гиорси.
- Олигофлексия : Oligoflexus .
- Acidithiobacillia : Acidithiobacillus ШоохШапз , Thermithiobacillus tepidarius
- Hydrogenophilalia : Hydrogenophilus thermoluteolus , Tepidiphilus margaritifer.

Трансформация
Трансформация , процесс, при котором генетический материал передается от бактерии к другой, описан по крайней мере у 30 видов «протеобактерий», распределенных по классам альфа, бета, гамма и эпсилон. Наиболее изученными «протеобактериями» в отношении естественной генетической трансформации являются важные с медицинской точки зрения патогены человека Neisseria gonorrhoeae (класс бета), Haemophilus influenzae (класс гамма) и Helicobacter pylori (класс эпсилон). Естественная генетическая трансформация — это половой процесс, включающий передачу ДНК от одной бактериальной клетки к другой через промежуточную среду и интеграцию донорной последовательности в геном реципиента. У патогенных «протеобактерий» трансформация, по-видимому, служит процессом репарации ДНК, который защищает ДНК патогена от атаки фагоцитарной защитой хозяина, использующей окислительные свободные радикалы .
Наиболее изученными «протеобактериями» в отношении естественной генетической трансформации являются важные с медицинской точки зрения патогены человека Neisseria gonorrhoeae (класс бета), Haemophilus influenzae (класс гамма) и Helicobacter pylori (класс эпсилон). Естественная генетическая трансформация — это половой процесс, включающий передачу ДНК от одной бактериальной клетки к другой через промежуточную среду и интеграцию донорной последовательности в геном реципиента. У патогенных «протеобактерий» трансформация, по-видимому, служит процессом репарации ДНК, который защищает ДНК патогена от атаки фагоцитарной защитой хозяина, использующей окислительные свободные радикалы .
использованная литература
внешние ссылки
Эшерихия — Escherichia — qaz.wiki
Кишечный является родом из грамотрицательных , не- споровой образующей, факультативно анаэробной , стержнеобразной бактерии из семейства Enterobacteriaceae . В тех видах которые являются обитателями желудочно — кишечного тракта от теплокровных животных, Escherichia видов обеспечить часть микробов полученного витамином К для хозяина. Некоторые виды Escherichia являются патогенными . Род назван в честь Теодора Эшериха , первооткрывателя Escherichia coli . Escherichia — это факультативные аэробы с аэробным и анаэробным ростом и оптимальной температурой 37 ° C. Escherichia обычно подвижны с помощью жгутиков , выделяют газ из ферментируемых углеводов и не декарбоксилируют лизин и не гидролизуют аргинин . Виды включают E. albertii , E. fergusonii , E. hermannii , E. senegalensis , E. marmotae и, прежде всего, модельный организм и клинически значимую E. coli . Shimwellia blattae ранее относилась к этому роду.
В тех видах которые являются обитателями желудочно — кишечного тракта от теплокровных животных, Escherichia видов обеспечить часть микробов полученного витамином К для хозяина. Некоторые виды Escherichia являются патогенными . Род назван в честь Теодора Эшериха , первооткрывателя Escherichia coli . Escherichia — это факультативные аэробы с аэробным и анаэробным ростом и оптимальной температурой 37 ° C. Escherichia обычно подвижны с помощью жгутиков , выделяют газ из ферментируемых углеводов и не декарбоксилируют лизин и не гидролизуют аргинин . Виды включают E. albertii , E. fergusonii , E. hermannii , E. senegalensis , E. marmotae и, прежде всего, модельный организм и клинически значимую E. coli . Shimwellia blattae ранее относилась к этому роду.
Патогенез
Хотя многие Escherichia являются комменсальными членами кишечной микробиоты , определенные штаммы некоторых видов, в первую очередь серотипы E. coli , являются патогенами человека и являются наиболее частой причиной инфекций мочевыводящих путей , важных источников желудочно-кишечных заболеваний, начиная от простых от диареи до состояний, похожих на дизентерию , а также от широкого спектра других патогенных состояний, в целом классифицируемых как эшерихиоз толстой кишки. В то время как E. coli ответственна за подавляющее большинство патогенеза, связанного с Escherichia , другие представители этого рода также участвуют в заболеваниях человека. Эшерихии связаны с нарушением баланса микробиоты нижних половых путей у женщин. Эти виды связаны с воспалением.
coli , являются патогенами человека и являются наиболее частой причиной инфекций мочевыводящих путей , важных источников желудочно-кишечных заболеваний, начиная от простых от диареи до состояний, похожих на дизентерию , а также от широкого спектра других патогенных состояний, в целом классифицируемых как эшерихиоз толстой кишки. В то время как E. coli ответственна за подавляющее большинство патогенеза, связанного с Escherichia , другие представители этого рода также участвуют в заболеваниях человека. Эшерихии связаны с нарушением баланса микробиоты нижних половых путей у женщин. Эти виды связаны с воспалением.
Смотрите также
Рекомендации
внешняя ссылка
- Геномы Escherichia и соответствующая информация в PATRIC , Ресурсном центре биоинформатики, финансируемом NIAID
Кишечная палочка
Микроорганизмы в пищевых продуктах присутствуют всегда. Патогенные микроорганизмы могут быть уничтожены благодаря обработке продуктов. Они могут стремительно размножаться из-за неправильной транспортировки, хранения, приготовления, или подачи. Увеличение количества этих микроорганизмов может привести не только к порче пищи, но и вызвать серьёзное отравление. Распространённый представитель пищевых патогенных бактерий — кишечная палочка. Кишечная палочка является частью микрофлоры кишечника человека и животных.
Патогенные микроорганизмы могут быть уничтожены благодаря обработке продуктов. Они могут стремительно размножаться из-за неправильной транспортировки, хранения, приготовления, или подачи. Увеличение количества этих микроорганизмов может привести не только к порче пищи, но и вызвать серьёзное отравление. Распространённый представитель пищевых патогенных бактерий — кишечная палочка. Кишечная палочка является частью микрофлоры кишечника человека и животных.
Некоторые виды кишечных палочек — патогенные для человека, то есть способны вызвать заболевание.
Бактерии группы кишечной палочки — универсальный показатель качества пищевых продуктов. Наличие кишечной палочки — показатель фекального загрязнения, особенно воды.
К сожалению, по внешнему виду, запаху или вкусу мы не сможем сказать, загрязнена ли пища кишечной палочкой (E. coli).
Кишечной палочкой могут быть обсеменены многие продукты, включая говядину, зелень, готовые к употреблению салаты, фрукты, сырое молоко и сырое тесто, нарезки колбас, сыров , особенно в условиях рынка, где не всегда обрабатывается аппарат для нарезки, мясорубки для приготовления фарша. Кишечная палочка активно размножается во время гниения продуктов.
Кишечная палочка активно размножается во время гниения продуктов.
Механизм передачи возбудителя фокально-оральный. Заражение происходит через пищу, воду, грязные руки.
Эта бактерия способна вырабатывать токсины (25 типов) и в зависимости от типа токсина, вырабатываемого кишечной палочкой, она обладают определенным действием.
Например, энтеротоксигенные E.coli имеют высокомолекулярный термолабильный токсин, который действует аналогично холерному, вызывая холероподобную диарею (гастроэнтериты у детей младшего возраста, диарею путешественников и др.).
Энтероинвазивные кишечные палочки вызывают профузную диарею с примесью крови и большим количеством лейкоцитов (аналогично дизентерие).
Энтеропатогенные E.coli вызыают водянистую диарею и выраженное обезвоживание.
Энтерогеморрагические кишечные палочки вызывают диарею с примесью крови.
Симптомы
Симптомы пищевого отравления отравления кишечной палочкой: боль в животе, тошнота, рвота, диарея более 20 раз в сутки, возможно с кровью. Температура тела обычно повышается незначительно или остаётся в норме.
Температура тела обычно повышается незначительно или остаётся в норме.
Кишечная палочка является наиболее распространенным патогеном, вызывающим менингит у новорождённых детей. Он имеет высокие показатели заболеваемости и смертности во всем мире.
В группе риска
-Взрослые в возрасте 65 лет и старше
-Дети младше 5 лет
-Люди с ослабленной иммунной системой, в том числе беременные
-Люди, которые путешествуют в определенные страны
Профилактика
Чтобы защитить себя от инфекций, вызванных кишечной палочкой, а также от других болезней пищевого происхождения, соблюдайте основные правила безопасности:
-Мойте руки, посуду и кухонные поверхности горячей мыльной водой до и после приготовления или приема пищи.
— Используйте отдельные разделочные доски для сырых продуктов и готовых
-Тщательно мойте фрукты и овощи, испотльзуйте щетку для овощей.
-Держите сырые продукты, особенно мясо и птицу, отдельно от готовых к употреблению продуктов.
— Охлаждайте или замораживайте скоропортящиеся продукты как можно быстрее.
— Избегайте непастеризованных соков, молочных продуктов.
— Не ешьте сырое тесто.
— Пейте бутилированную воду.
-Тщательно прожаривайте мясо.
Characterization of Membrane Transporters by Heterologous Expression in E. coli and Production of Membrane Vesicles
Основные шаги в этом протоколе обобщены живописно на рисунке 1. Короче говоря, клетки кишечной палочки, недостающие во всех обменных пунктах полиамина и выражающие AtBAT1, культивируются, центрифугируются, промываются буфером и подвергаются клеточному лисису с помощью французской прессы. Lysis, как правило, производят пузырьки, которые в основном наизнанку и ловушки буфера за пределами клеток. Клеточный мусор удаляется центрифугированием, и для сбора мембранных гранул используется второй этап ультрацентрифирования. Мембранные гранулы перевешивается в буфере Tris-Maleate pH 5. 2 и хранятся при -80 градусах Цельсия. Транспортные анализы проводятся при 12 градусах Цельсия, что оказалось оптимальным для поддержания мембранной стабильности. Анализы инициируются добавлением радиомаркированного субстрата и сдвигом в рН буферной подвески пузырьков до рН 8.0. После 1 мин, ледяной асси буфер с немаркированными субстратами добавляется, чтобы остановить поглощение радиомаркировки в пузырьки. Радиомаркированные пузырьки застревают фильтрацией через мембраны нитроцеллюлозы. Мембраны передаются в сцинтилляционные флаконы, а радиометка на мембранах определяется подсчетом жидкого сцинтилляции.
2 и хранятся при -80 градусах Цельсия. Транспортные анализы проводятся при 12 градусах Цельсия, что оказалось оптимальным для поддержания мембранной стабильности. Анализы инициируются добавлением радиомаркированного субстрата и сдвигом в рН буферной подвески пузырьков до рН 8.0. После 1 мин, ледяной асси буфер с немаркированными субстратами добавляется, чтобы остановить поглощение радиомаркировки в пузырьки. Радиомаркированные пузырьки застревают фильтрацией через мембраны нитроцеллюлозы. Мембраны передаются в сцинтилляционные флаконы, а радиометка на мембранах определяется подсчетом жидкого сцинтилляции.
Западная поместье используется для проверки того, что AtBAT1 трансумбит в пузырьки(рисунок 2). Зондирование пятно с анти-Его C-терминал антитела показали синтез агенство белка примерно 72,3 kDa(Рисунок 2, переулок 2). Переваривание пузырьков до SDS-PAGE привело к диминитии, но не полная потеря сигнала зонда(рисунок 2,полоса 3). Уменьшение сигнала зонда в результате карбоксипептидазы А говорит о том, что большинство остатков C-терминала находятся на внешней стороне пузырьков.
Уменьшение сигнала зонда в результате карбоксипептидазы А говорит о том, что большинство остатков C-терминала находятся на внешней стороне пузырьков.
В этой системе асссея пузырьки подвешиваются в буфере на рН 5.2, так что рН внутри пузырьков уравновешивается буфером. Транспортировка радиомаркированного субстрата в пузырьки на рН 5.2 инициируется приостановкой пузырьков в буфере pH 8.0, создавая тем самым градиент рН рН 2.8 по всей мембране. При 12 градусах Цельсия поглощение радиомаркированного спермидина пузырьками было самым высоким на 1 мин, и оставалось линейным более 3 мин(рисунок 3A). Таким образом, время инкубации для транспортного ассеа было зафиксировано на 1 мин. Для учета неспецифических связывания радиомаркировки пузырьки инкубировались при 0 градусах Цельсия при наличии радиомаркированного субстрата в течение одной минуты, и эти подсчеты вычитались из поглощения ладиоlabeli при более высоких температурах.
На рисунке 3B показано поглощение радиомаркированного спермидина в пузырьки через одну минуту. Не было никакого чистого поглощения изотопов мембранными пузырьками, которые были подготовлены и хранятся при рН 8.0, так как не было градиента протона через везикулм. Чтобы продемонстрировать эффект рассеивания искусственного градиента протона, мембранные пузырьки были инкубированы в pH 8.0 буфер в течение 10 минут до добавления маркированного субстрата25. В этих условиях было показано минимальное поглощение радиомаркированного субстрата. Поглощение радиомаркированного спермидина было также минимальным в пузырьках, подготовленных с e. coli клеток, недостаточно в полиаминных обменников CadB и PotE. Взятые вместе, эти результаты показывают, что протон инициативе поглощения спермидина было связано с белком BAT1 (Рисунок 3A,B).
 within-page=»1″>Для определения специфики субстрата белка значения Km были рассчитаны путем измерения поглощения радиомаркированного субстрата в концентрациях 10, 25, 50, 100, 250 и 500 мкм. Kм для спермидина, гектарскина и аргинина были 55 и 12 мкм, 85 и 20 мкм и 1,4 и 0,5 мм соответственно, что указывает на то, что этот белок является высокой сродством полиамина и аргинина обменник(рисунок 4).
within-page=»1″>Для определения специфики субстрата белка значения Km были рассчитаны путем измерения поглощения радиомаркированного субстрата в концентрациях 10, 25, 50, 100, 250 и 500 мкм. Kм для спермидина, гектарскина и аргинина были 55 и 12 мкм, 85 и 20 мкм и 1,4 и 0,5 мм соответственно, что указывает на то, что этот белок является высокой сродством полиамина и аргинина обменник(рисунок 4).Сродство транспортера к конкретному субстрату также может быть определено косвенно с помощью конкурсных анализов. Здесь мы использовали два метода для оценки конкуренции между двумя субстратами. В первом методе поглощение 50 мкм радиомаркированный спермидин измерялся в присутствии увеличения концентраций немаркированного конкурирующего субстрата(рисунок 5А). Во втором методе, очевидно Kм для спермидина был рассчитан путем измерения поглощения увеличения концентрации радиомаркированных спермидин в присутствии 100 мКм немаркированных конкурирующих субстрата(Рисунок 5B). Конкурсные анализы показали, что ГАМК является конкурентоспособным ингибитором спермидина с Kм, приложение 164 и 15 мкм(рисунок 5A,B). Кроме того, измерение поглощения 50 мКМ радиомаркированных спермидин в присутствии различных концентраций различных аминокислот показали, что AtBAT1 также способен транспортировать глутамат и аланин в концентрациях ММ (Рисунок 6).
Конкурсные анализы показали, что ГАМК является конкурентоспособным ингибитором спермидина с Kм, приложение 164 и 15 мкм(рисунок 5A,B). Кроме того, измерение поглощения 50 мКМ радиомаркированных спермидин в присутствии различных концентраций различных аминокислот показали, что AtBAT1 также способен транспортировать глутамат и аланин в концентрациях ММ (Рисунок 6).
Рисунок 1: Схематическое представление метода. (A) Схематическое представление с изложением ключевых шагов в подготовке и очищении мембранных пузырьков от кишечной палочки. (B) Схематическое представление с изложением ключевых шагов в транспортном ассие мембранных препаратов везикля с использованием радиомаркированных субстратов. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
Рисунок 2: Западная поместье, показывающая выражение AtBAt1 в очищенных пузырьках. Полосы были визуализированы с использованием хрена peroxidase сопряжены анти-His (C-термин)-HRP антитела. Переулок 1, Запятнанная белковая лестница. Лейн 2, Очищенные пузырьки, выражающие AtBAT1.1, показывающие полосу ожидаемого размера белка синтеза. Лейн 3, очищенные пузырьки, выражающие AtBAT1.1, были предварительно обработаны карбоксипептидазой А до электрофореза СДС и западного промотирования. Эквивалентное количество пузырьков (белка) было добавлено к каждой полосе. Снижение окрашивания указывает на то, что C-терминал белка в большинстве пузырьков деградирует путем переваривания протеазы. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
Полосы были визуализированы с использованием хрена peroxidase сопряжены анти-His (C-термин)-HRP антитела. Переулок 1, Запятнанная белковая лестница. Лейн 2, Очищенные пузырьки, выражающие AtBAT1.1, показывающие полосу ожидаемого размера белка синтеза. Лейн 3, очищенные пузырьки, выражающие AtBAT1.1, были предварительно обработаны карбоксипептидазой А до электрофореза СДС и западного промотирования. Эквивалентное количество пузырьков (белка) было добавлено к каждой полосе. Снижение окрашивания указывает на то, что C-терминал белка в большинстве пузырьков деградирует путем переваривания протеазы. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
Рисунок 3: Транспортная активность пузырьков, показывающих влияние экспрессии белка BAT1 и важность градиента рН. (A) Время зависимых поглощения 3H помечены спермидин в пузырьков, выражающих BAT1 с внутренним рН 5,2 и введены в буфер на рН 8,0. В контрольном ассиде, пузырьки были добавлены в буфер ассеа на рН 8.0, 10 минут до добавления 3H помечены спермидин, чтобы позволить рассеивание градиента протона. Затем поглощение радиомаркировки в пузырьки оценивалось с интервалом в 1 минуту. (B) Поглощение 3H помечены спермидин в присутствии градиента протона (внутренний рН 5,2), при отсутствии градиента протона (внутренний рН 8), в пузырьки добавил к ассеа решение 10 мин до добавления радиомаркированных спермидина и в пузырьки из E. coli мутантных клеток, не выражающихся BAT1. Поглощение пузырьков контролировалось в течение 1 мин. Все значения представлены как средние sE пяти репликатов. Анализ данных был выполнен с использованием t-теста студента и q указывает на значительное отличие от элемента управления(p значение 0,05). Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
В контрольном ассиде, пузырьки были добавлены в буфер ассеа на рН 8.0, 10 минут до добавления 3H помечены спермидин, чтобы позволить рассеивание градиента протона. Затем поглощение радиомаркировки в пузырьки оценивалось с интервалом в 1 минуту. (B) Поглощение 3H помечены спермидин в присутствии градиента протона (внутренний рН 5,2), при отсутствии градиента протона (внутренний рН 8), в пузырьки добавил к ассеа решение 10 мин до добавления радиомаркированных спермидина и в пузырьки из E. coli мутантных клеток, не выражающихся BAT1. Поглощение пузырьков контролировалось в течение 1 мин. Все значения представлены как средние sE пяти репликатов. Анализ данных был выполнен с использованием t-теста студента и q указывает на значительное отличие от элемента управления(p значение 0,05). Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
Рисунок 4: In vitro анализы полиамина и аргинина транспортной активности BAT1. (A) Значения Km для спермидина и поглощения путрешкина 55 и 12 мкм и 85 и 32 м соответственно. (B) Kм для поглощения аргинина составляет 1,4 и 0,5 мм. Все значения представлены как средние sE пяти репликатов. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
(A) Значения Km для спермидина и поглощения путрешкина 55 и 12 мкм и 85 и 32 м соответственно. (B) Kм для поглощения аргинина составляет 1,4 и 0,5 мм. Все значения представлены как средние sE пяти репликатов. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
Рисунок 5: ГАМК является конкурентоспособным ингибитором спермидина транспорта BAT1. (A) Поглощение 3H помечены спермидин пузырьков, выражающих AtBAT1.1 был значительно сокращен в присутствии 100 мкм или 500 ММ ГАМК. (B) Явный Kм для поглощения спермидина BAT1.1 был увеличен до 164 и 20 мкм в присутствии 100 ММ ГАМК. Все значения представлены как средние sE пяти репликатов. Анализ данных был выполнен с использованием t-теста студента и q указывает на значительное отличие от элемента управления(p значение 0,05). Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
Рисунок 6: Глутамат и аланин являются конкурентоспособными ингибиторами спермидина transportby BAT1. Поглощение спермидина было значительно уменьшено при наличии 1 мМ немаркированного глутамата и 1,5 мМ немаркированного аланина. Все значения представлены как средние sE пяти репликатов. Анализ данных был выполнен с использованием t-теста студента и q указывает на значительное отличие от элемента управления(p значение 0,05). Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
Subscription Required. Please recommend JoVE to your librarian.
Паспортов безопасности патогенов: Инфекционные вещества — Escherichia coli, энтеропатогенные
ПАСПОРТ БЕЗОПАСНОСТИ ОТ ПАТОГЕНОВ — ИНФЕКЦИОННЫЕ ВЕЩЕСТВА
РАЗДЕЛ I — ИНФЕКЦИОННЫЙ АГЕНТ
НАЗВАНИЕ : Escherichia coli , энтеропатогенный
СИНОНИМ ИЛИ ПЕРЕКРЕСТНАЯ ССЫЛКА : EPEC Сноска 1, Сноска 2 , патогенный кишечник E coli Сноска 3 , острая и затяжная детская диарея Сноска 4 .
ХАРАКТЕРИСТИКИ : Энтеропатогенные Escherichia coli (EPEC) относятся к семейству Enterobacteriaceae Footnote 2 . Бактерии грамотрицательные, палочковидные, неспорообразующие, подвижные с перитрихозными жгутиками или неподвижные, растут на агаре МакКонки (колонии имеют диаметр 2–3 мм, красные или бесцветные). Footnote 5 . Они могут расти в аэробных и анаэробных условиях и не производят энтеротоксины. Footnote 1 .
РАЗДЕЛ II — ИДЕНТИФИКАЦИЯ ОПАСНОСТИ
ПАТОГЕННОСТЬ / ТОКСИЧНОСТЬ : EPEC вызывает острую, обильную, водянистую диарею, которая редко становится стойкой. Footnote 1 .Стул обычно не кровавый, слизистый или дизентерийный. Может присутствовать субфебрильная температура с тошнотой и рвотой Footnote 1, Footnote 6 . EPEC характеризуется образованием прикрепляющихся и смещающихся поражений в пораженных участках кишечника. Footnote 7 . В настоящее время распознаются две группы EPEC: типичная EPEC и атипичная EPEC. Типичный EPEC, основная причина детской диареи в развивающихся странах, редко встречается в промышленно развитых странах, где атипичный EPEC, по-видимому, является более важной причиной диареи. Footnote 8 .Типичные и атипичные EPEC также различаются по генетическим характеристикам, серотипам и свойствам вирулентности. Атипичный EPEC более тесно связан с E.coli, продуцирующей токсин Shiga (STEC), и, как и STEC, эти штаммы, по-видимому, являются появляющимися патогенами. Footnote 8 . Атипичные штаммы EPEC могут быть менее вирулентными, чем типичные. Одной из причин может быть отсутствие плазмиды фактора адгезии EPEC (EAF) Footnote 8 .
Типичный EPEC, основная причина детской диареи в развивающихся странах, редко встречается в промышленно развитых странах, где атипичный EPEC, по-видимому, является более важной причиной диареи. Footnote 8 .Типичные и атипичные EPEC также различаются по генетическим характеристикам, серотипам и свойствам вирулентности. Атипичный EPEC более тесно связан с E.coli, продуцирующей токсин Shiga (STEC), и, как и STEC, эти штаммы, по-видимому, являются появляющимися патогенами. Footnote 8 . Атипичные штаммы EPEC могут быть менее вирулентными, чем типичные. Одной из причин может быть отсутствие плазмиды фактора адгезии EPEC (EAF) Footnote 8 .
ЭПИДЕМИОЛОГИЯ : Типичный EPEC в первую очередь вызывает заболевание у новорожденных и детей младшего возраста, при этом большинство случаев встречается у детей младше 2 лет и особенно у детей младше 6 месяцев. Footnote 1 .Заболевание может возникнуть у взрослых при приеме внутрь достаточно большого количества инокулята. Вспышки произошли в педиатрических палатах, яслях и детских садах, а также среди взрослых, которые потребляли зараженную пищу из буфета. В развивающихся странах EPEC широко распространены и являются важной причиной детских диарейных заболеваний и смертей, связанных с обезвоживанием. Исследования, проведенные в Бразилии, Мексике и Южной Африке, показали, что 30–40% случаев детской диареи могут быть отнесены к EPEC Footnote 6 . Атипичные EPEC распространены как в развитых, так и в развивающихся странах.По всей видимости, они вызывают заболевания в более широком диапазоне возрастов и были связаны со вспышками в развитых странах. Footnote 9 . Однако энтеропатогенность и роль некоторых атипичных штаммов EPEC являются спорными. Footnote 10 . По крайней мере, два исследования «случай-контроль» не показали статистической разницы в уровне инфицирования между случаями и подобранной контрольной группой, что позволяет предположить, что они могут быть частью нормальной флоры человека.
Вспышки произошли в педиатрических палатах, яслях и детских садах, а также среди взрослых, которые потребляли зараженную пищу из буфета. В развивающихся странах EPEC широко распространены и являются важной причиной детских диарейных заболеваний и смертей, связанных с обезвоживанием. Исследования, проведенные в Бразилии, Мексике и Южной Африке, показали, что 30–40% случаев детской диареи могут быть отнесены к EPEC Footnote 6 . Атипичные EPEC распространены как в развитых, так и в развивающихся странах.По всей видимости, они вызывают заболевания в более широком диапазоне возрастов и были связаны со вспышками в развитых странах. Footnote 9 . Однако энтеропатогенность и роль некоторых атипичных штаммов EPEC являются спорными. Footnote 10 . По крайней мере, два исследования «случай-контроль» не показали статистической разницы в уровне инфицирования между случаями и подобранной контрольной группой, что позволяет предположить, что они могут быть частью нормальной флоры человека. Footnote 11-Footnote 13 . Вероятно, что данные о EPEC и, в частности, об атипичных EPEC значительно занижены.
Footnote 11-Footnote 13 . Вероятно, что данные о EPEC и, в частности, об атипичных EPEC значительно занижены.
ДИАПАЗОН ПРИЕМНИКОВ : Люди Сноска 1 и животные, включая крупный рогатый скот и лошадей Сноска 14 .
ИНФЕКЦИОННАЯ ДОЗА : Инфекционная доза EPEC для здоровых взрослых оценивается в 10 6 организмов Footnote 15 .
СПОСОБ ПЕРЕДАЧИ : Загрязненная пища, вода и фомиты служат носителями фекальной / оральной передачи EPEC Footnote 1, Footnote 16 .
ПЕРИОД ИНКУБАЦИИ : Инкубационный период составляет от 6 до 48 часов Сноска 17 .
КОММУНИКАЦИЯ : Может передаваться при прямом контакте с инфицированными людьми Footnote 14 .
РАЗДЕЛ III — РАСПРОСТРАНЕНИЕ
РЕЗЕРВУАР : Типичный EPEC: Люди Сноска 18 . Атипичный EPEC: собаки, кошки, крупный рогатый скот, овцы, кролики, обезьяны и люди Footnote 8, Footnote 9 .
ЗООНОЗ : Да. Может передаваться при контакте с инфицированным скотом, собаками, кошками, овцами, кроликами и лошадьми. Footnote 19 .
ВЕКТОРОВ : Нет.
РАЗДЕЛ IV — УСТОЙЧИВОСТЬ И ЖЕСТКОСТЬ
ЛЕКАРСТВЕННАЯ УСТОЙЧИВОСТЬ / УСТОЙЧИВОСТЬ : Чувствительность к карбапенему, фосфомицин-трометанол, нитрофурантоин и бычий аполактоферрин. E. coli может быть устойчивым к хлорамфениколу, β-лактамам, налидиксовой кислоте, ампициллину и ципрофлоксацину. Фторхинолоны, такие как ципрофлоксацин, увеличивают выработку токсинов Footnote 3, Footnote 20 .
УСТОЙЧИВОСТЬ К ДЕЗИНФЕКЦИОННЫМ СРЕДСТВАМ : Чувствительность к комбинации 2,2-дибром-2-цианоацетамида (DBA) с йодидом натрия (20:80 частей), йодом, 2% глутаральдегидом, четвертичным аммонием (20 ° C, 0.5 мин), гипохлорит (0,525%, 20 ° C, 0,5 мин), фенольные смолы (20 ° C, 0,5 мин) и этиловый спирт (70%, 20 ° C, 0,5 мин) Footnote 21-Footnote 23 .
ФИЗИЧЕСКАЯ ИНАКТИВАЦИЯ : Озон может инактивировать E. coli Footnote 24 . E. coli также чувствительна к термической обработке, особенно при температуре 70 ° C и выше. Footnote 19, Footnote 25 .
ВЫЖИВАНИЕ ВНЕ ХОЗЯЙСТВА : E. coli может выжить в течение 1.От 5 часов до 16 месяцев на сухих неодушевленных поверхностях Footnote 26 .
РАЗДЕЛ V — ПЕРВАЯ ПОМОЩЬ / МЕДИЦИНСКАЯ
НАБЛЮДЕНИЕ : Следите за симптомами. Культивирование стула — распространенный метод, используемый для идентификации E. coli Footnote 27 . ДНК-зонды и методы, такие как ПЦР, можно применять непосредственно к клиническим образцам и продуктам питания. Footnote 3 . Как типичные, так и атипичные EPEC чаще всего идентифицируются путем обнаружения гена eae , кодирующего белок intimin.Наличие гена eae и демонстрация отсутствия гена веротоксина (энтеротоксина) абсолютно необходимы для молекулярной идентификации EPEC Footnote 1 . Чтобы идентифицировать типичный EPEC, можно использовать методы гибридизации ПЦР или ДНК для обнаружения плазмиды EAF и гена, кодирующего bfp Footnote 1, Footnote 3 . Дополнительные тесты, которые могут быть выполнены в некоторых ситуациях, — это тест на флуоресцентное окрашивание актина и тесты на приверженность, показывающие локализованную приверженность (LA).
Чтобы идентифицировать типичный EPEC, можно использовать методы гибридизации ПЦР или ДНК для обнаружения плазмиды EAF и гена, кодирующего bfp Footnote 1, Footnote 3 . Дополнительные тесты, которые могут быть выполнены в некоторых ситуациях, — это тест на флуоресцентное окрашивание актина и тесты на приверженность, показывающие локализованную приверженность (LA).
Примечание. Не все методы диагностики доступны во всех странах.
ПЕРВАЯ ПОМОЩЬ : Лечение триметопримом / сульфаметоксазолом (TMP-SMX) или хинолонами сокращает продолжительность диареи. Footnote 28 . Лечение потери жидкости и электролитов обычно достигается за счет пероральной регидратации. Footnote 19 . Рекомендуется использовать раствор солей для пероральной регидратации (ПРС) Всемирной организации здравоохранения. Footnote 1 .Внутривенная регидратация может быть необходима младенцам, людям с чрезмерной рвотой или людям с тяжелым обезвоживанием. Субсалицилат висмута может уменьшить количество диареи и продолжительность заболевания. Антимикробная терапия, как правило, не показана из-за того, что большинство из этих заболеваний носит ограниченный характер.
Субсалицилат висмута может уменьшить количество диареи и продолжительность заболевания. Антимикробная терапия, как правило, не показана из-за того, что большинство из этих заболеваний носит ограниченный характер.
ИММУНИЗАЦИЯ : В настоящее время нет одобренных для использования человеком вакцин против диареи E. coli Footnote 1 .
PROPHYLAXIS : TMP-SMX рекомендуется для краткосрочного использования (<2 недель) для людей с высоким риском заболевания. Footnote 28 .Субсалицилат висмута оказывает некоторое профилактическое действие, но не должен использоваться вместо других профилактических мер. Footnote 1 .
РАЗДЕЛ VI — ЛАБОРАТОРНЫЕ ОПАСНОСТИ
ЛАБОРАТОРНЫЕ ИНФЕКЦИИ : было зарегистрировано 12 случаев лабораторных инфекций, вызванных E. coli , большинство из которых были вызваны энтерогеморрагической E.coli (EHEC) Footnote 29 .
ИСТОЧНИКИ / ОБРАЗЦЫ : Стул и фекально загрязненный материал Сноска 1, Сноска 6, Сноска 16 .
ОСНОВНАЯ ОПАСНОСТЬ : Проглатывание Сноска 29 .
ОСОБАЯ ОПАСНОСТЬ : Нет.
РАЗДЕЛ VII — КОНТРОЛЬ ВОЗДЕЙСТВИЯ / ЛИЧНАЯ ЗАЩИТА
КЛАССИФИКАЦИЯ ГРУПП РИСКА : Группа риска 2 Сноска 30 .
ТРЕБОВАНИЯ К СОДЕРЖАНИЮ : Помещения, оборудование и методы работы уровня сдерживания 2 для работы с инфекционными или потенциально инфекционными материалами, животными или культурами Сноска 31 .
ЗАЩИТНАЯ ОДЕЖДА : Лабораторный халат. Перчатки при неизбежном прямом контакте кожи с инфицированными материалами или животными. Средства защиты глаз должны использоваться там, где существует известный или потенциальный риск воздействия брызг.
ДРУГИЕ МЕРЫ ПРЕДОСТОРОЖНОСТИ : Все процедуры, которые могут привести к образованию аэрозолей, или связаны с высокими концентрациями или большими объемами, должны проводиться в шкафу биологической безопасности (BSC). Footnote 31 . Использование игл, шприцев и других острых предметов должно быть строго ограничено.Дополнительные меры предосторожности следует учитывать при работе с животными или крупномасштабной деятельности.
Footnote 31 . Использование игл, шприцев и других острых предметов должно быть строго ограничено.Дополнительные меры предосторожности следует учитывать при работе с животными или крупномасштабной деятельности.
РАЗДЕЛ VIII — ОБРАЩЕНИЕ И ХРАНЕНИЕ
РАЗЛИВЫ : Дайте аэрозолям осесть и, надев защитную одежду, аккуратно накройте разлив бумажными полотенцами и нанесите соответствующее дезинфицирующее средство, начиная с периметра и двигаясь к центру. Перед очисткой дайте достаточно времени для контакта. Footnote 31 .
УТИЛИЗАЦИЯ : Обеззараживайте все отходы, которые содержат инфекционный организм или контактировали с ним, с помощью автоклава, химической дезинфекции, гамма-облучения или сжигания перед утилизацией. Footnote 31 .
ХРАНЕНИЕ : Инфекционный агент должен храниться в герметичных контейнерах, имеющих соответствующую маркировку Сноска 31 .
РАЗДЕЛ IX — НОРМАТИВНАЯ И ДРУГАЯ ИНФОРМАЦИЯ
НОРМАТИВНАЯ ИНФОРМАЦИЯ : Импорт, транспортировка и использование патогенных микроорганизмов в Канаде регулируется многими регулирующими органами, включая Агентство общественного здравоохранения Канады, Министерство здравоохранения Канады, Канадское агентство по инспекции пищевых продуктов, Министерство окружающей среды Канады и Министерство транспорта Канады. Пользователи несут ответственность за соблюдение всех соответствующих законов, постановлений, руководств и стандартов.
Пользователи несут ответственность за соблюдение всех соответствующих законов, постановлений, руководств и стандартов.
ОБНОВЛЕНО : декабрь 2011 г.
ПОДГОТОВЛЕНО : Управление по регулированию патогенов, Агентство общественного здравоохранения Канады
Хотя информация, мнения и рекомендации, содержащиеся в этом Паспорте безопасности патогенов, собраны из источников, которые считаются надежными, мы не несем ответственности за точность, достаточность или надежность, а также за любые потери или травмы, возникшие в результате использования информации.Часто обнаруживаются новые опасности, и эта информация может быть не полностью актуальной.
Авторские права ©
Агентство общественного здравоохранения Канады, 2011 г.
Канада
Escherichia coli — обзор
Общие характеристики
Escherichia coli является членом семейства Enterobacteriaceae, которое включает грамотрицательные, факультативно анаэробные палочковидные бактерии (обладающие как ферментативным, так и респираторным метаболизмом), которые не производят фермент оксидаза. Клетки Escherichia coli обычно имеют ширину 1,1–1,5 мкм, длину 2–6 мкм и представляют собой отдельные прямые стержни. Они могут быть как подвижными, так и неподвижными, и, когда они подвижны, образуют латеральные, а не полярные жгутики. Помимо жгутиков, многие штаммы продуцируют другие придатки, такие как фимбрии или пили, которые представляют собой белковые структуры (или придатки или волокна), которые выходят наружу от бактериальной поверхности и играют роль в прикреплении к поверхностям, включая другие клетки или ткани хозяина.
Клетки Escherichia coli обычно имеют ширину 1,1–1,5 мкм, длину 2–6 мкм и представляют собой отдельные прямые стержни. Они могут быть как подвижными, так и неподвижными, и, когда они подвижны, образуют латеральные, а не полярные жгутики. Помимо жгутиков, многие штаммы продуцируют другие придатки, такие как фимбрии или пили, которые представляют собой белковые структуры (или придатки или волокна), которые выходят наружу от бактериальной поверхности и играют роль в прикреплении к поверхностям, включая другие клетки или ткани хозяина.
Escherichia coli имеет штамм-специфические O-липополисахаридные антигены на своей клеточной стенке (в настоящее время распознаются по крайней мере 188 O-антигенов) и антигены жгутиков или H, если они присутствуют (распознаются по крайней мере 53 H-типа).Существует также множество различных капсульных полисахаридных (K) антигенов. Escherichia coli серотипированы на основе комбинации антигенов O, H и K, хотя обычно перечислены только типы O и H, например, E. coli O157: H7. Серотипирование E. coli вместе с геномом, вирулентностью и типированием фага является полезным эпидемиологическим инструментом. Полногеномное секвенирование — это метод, который все чаще используется для типирования E. coli и идентификации генов вирулентности.Непатогенные последовательности генома E. coli состоят из 4,6 миллиона оснований, в то время как патогенные изоляты имеют более крупные геномы — около 5,4 миллиона оснований. Такая геномная информация обеспечивает основу для понимания взаимосвязей между различными патогенными группами E. coli , способности изолятов вызывать заболевание и способности передавать генетический материал, который может привести к появлению новых или более вирулентных типов E Coli .
coli O157: H7. Серотипирование E. coli вместе с геномом, вирулентностью и типированием фага является полезным эпидемиологическим инструментом. Полногеномное секвенирование — это метод, который все чаще используется для типирования E. coli и идентификации генов вирулентности.Непатогенные последовательности генома E. coli состоят из 4,6 миллиона оснований, в то время как патогенные изоляты имеют более крупные геномы — около 5,4 миллиона оснований. Такая геномная информация обеспечивает основу для понимания взаимосвязей между различными патогенными группами E. coli , способности изолятов вызывать заболевание и способности передавать генетический материал, который может привести к появлению новых или более вирулентных типов E Coli .
Escherichia coli тесно связана с Shigella spp., хотя Shigella имеет тенденцию быть менее биохимически активным, чем большинство штаммов E. coli . Shigella и E. coli могут рассматриваться как принадлежащие к одному роду на основании генетического родства, но исторически эти два вида оставались отдельными, чтобы избежать путаницы в медицинском диагнозе.
coli могут рассматриваться как принадлежащие к одному роду на основании генетического родства, но исторически эти два вида оставались отдельными, чтобы избежать путаницы в медицинском диагнозе.
Краткий обзор Escherichia coli O157: H7 и его плазмиды O157
J Microbiol Biotechnol. Авторская рукопись; доступно в PMC 2013 6 мая.
Опубликован в окончательной редакции как:
J Microbiol Biotechnol.2010 Янв; 20 (1): 5–14.
PMCID: PMC3645889
NIHMSID: NIHMS464337
Ji Youn Lim
1 Кафедра микробиологии, молекулярной биологии и биохимии, Университет Айдахо, Москва, Айдахо 83844, США W. Yang
Jang 2 Группа передовых кадров и исследований в области медицины (BK21), Медицинский факультет Университета Конкук, Сеул 143-701, Корея Кэролайн Дж. Ховде
1 Департамент микробиологии, молекулярной биологии и биохимии, Университет Айдахо , Москва, Айдахо 83844, У.SA
1 Кафедра микробиологии, молекулярной биологии и биохимии, Университет Айдахо, Москва, Айдахо 83844, США
2 Группа передовых человеческих ресурсов и исследований в области медицины (BK21), Медицинский факультет Университета Конкук, Сеул 143-701, Корея
См. Другие статьи в PMC, в которых цитируется опубликованная статья. Реферат
Энтерогеморрагический Escherichia coli O157: H7 — основной патоген пищевого происхождения, вызывающий тяжелые заболевания людей во всем мире.Здоровый крупный рогатый скот является резервуаром E. coli O157: H7, а продукты питания крупного рогатого скота и свежие продукты, загрязненные бычьими отходами, являются наиболее частыми источниками вспышек заболеваний в Соединенных Штатах . E. coli O157: H7 также хорошо выживает в окружающей среде. Способность вызывать заболевание человека, колонизировать желудочно-кишечный тракт крупного рогатого скота и выживать в окружающей среде требует, чтобы E. coli O157: H7 адаптировались к широкому спектру условий. Три основных фактора вирулентности E.coli O157: H7, включая токсины Шига, продукты острова патогенности, называемого локусом сглаживания энтероцитов, и продукты F-подобной плазмиды pO157. Среди этих факторов вирулентности роль pO157 изучена меньше всего. В этом обзоре представлен обзор платы E. coli O157: H7 с акцентом на pO157.
Ключевые слова: E. coli O157: H7, pO157
Escherichia coli O157: H7 Escherichia coli
Escherichia coli ( E.coli ) — грамотрицательная палочковидная факультативная анаэробная бактерия. Этот микроорганизм был впервые описан Теодором Эшерихом в 1885 году. Большинство штаммов E. coli безвредно колонизируют желудочно-кишечный тракт людей и животных как нормальная флора. Однако есть некоторые штаммы, которые превратились в патогенную E. coli , приобретая факторы вирулентности через плазмиды, транспозоны, бактериофаги и / или островки патогенности. Эти патогенные E. coli могут быть классифицированы на основе серогрупп, механизмов патогенности, клинических симптомов или факторов вирулентности [33, 47].Среди них энтерогеморрагическая E. coli (EHEC) определяется как патогенные штаммы E. coli , которые продуцируют токсины Шига (Stxs) и вызывают геморрагический колит (HC) и опасные для жизни последствия гемолитико-уремического синдрома (HUS) у людей. . Некоторые серотипы в EHEC часто связаны с заболеваниями человека, такими как O26: h21, O91: h31, O111: H8, O157: NM и O157: H7 [44, 51]. E. coli O157: H7 — серотип EHEC, наиболее часто выделяемый у больных в США, Японии и Великобритании, и он является предметом настоящего обзора.
История
Серотип EHEC O157: H7 был впервые признан в 1982 г. как патоген человека, связанный со вспышками кровавой диареи в Орегоне и Мичигане, США [57, 71], а также был связан со спорадическими случаями HUS в 1983 г. [34] . С тех пор в США было зарегистрировано множество вспышек, связанных с EHEC, и E. coli O157: H7 стала одним из наиболее важных патогенов пищевого происхождения.
Распространенность и экономические затраты
По оценкам Центров по контролю и профилактике заболеваний (CDC), E. coli O157: инфекции H7 вызывают 73 000 заболеваний, 2200 госпитализаций и 60 смертей ежегодно в США [43]. Данные эпиднадзора за вспышками, предоставленные CDC, показали, что инфекций E. coli O157: H7 сокращаются после пика в 1999 г. Однако крупные вспышки и спорадические случаи продолжают иметь место. Ежегодные затраты на заболевание, вызванное инфекциями E. coli O157: H7, составили 405 миллионов долларов, включая потерю производительности, медицинское обслуживание и преждевременную смерть [21]. Высокая стоимость болезни требует дополнительных усилий по борьбе с этим возбудителем.
Выделение и идентификация
E. coli O157: H7 экспрессирует соматический (O) антиген 157 и антиген жгутиков (H) 7. E. coli O157: H7 обладает уникальными свойствами замедленной ферментации D-сорбита (> 24 з) и неспособность продуцировать β-глюкуронидазу, которая может гидролизовать синтетическую молекулу, 4-метил-умбеллиферил-D-глюкуронид (MUG) [68]. Таким образом, агар Sorbitol MacConkey (SMAC) с добавлением MUG был использован для обнаружения E. coli O157: H7. Для увеличения селективности на E.coli O157: H7, цефиксим, теллурит калия и ванкомицин были добавлены в чашки с агаром SMAC для подавления другой грамотрицательной флоры. Серотипы O157 и H7 могут быть дополнительно подтверждены коммерчески доступным анализом латексной агглютинации.
Геномная организация
Размер хромосомы E. coli O157: H7 составляет 5,5 Мб. Этот геном включает в себя последовательность из 4,1 млн п.н., законсервированную во всех штаммах E. coli . Остальные специфичны для E. coli O157: H7 [53].Кроме того, сравнение генома E. coli O157: H7 с непатогенным E. coli K12 показывает, что 0,53 МБ ДНК отсутствует для E. coli O157: H7, что позволяет предположить, что сокращение генома также сыграло роль в E coli O157: эволюция H7 [17, 53]. Большинство последовательностей ДНК E. coli O157: H7 (1,4 МБ) представляют собой горизонтально переносимые чужеродные ДНК, такие как профаг и профагоподобные элементы. E. coli O157: H7 содержит 463 фаго-ассоциированных гена по сравнению с 29 в E.coli K-12 [72]. Изменение содержания G + C является одним из признаков того, что геномная область была получена путем горизонтального переноса, и Putonti et al. [55] подсчитал, что по крайней мере 53 различных вида внесли вклад в эти уникальные последовательности в E. coli O157: H7. Связанные с вирулентностью гены между двумя секвенированными штаммами E. coli O157: H7 почти идентичны (99%). Очевидно, что как приобретение, так и потеря ДНК сыграли важную роль в эволюции патогенеза E.coli O157: H7.
Evolution
Несколько сравнительных и эпидемиологических исследований показывают, что E. coli O157: H7, возможно, произошли от нетоксигенного и менее вирулентного штамма E. coli O55: H7 [72]. E. coli O15: H7 возник в результате четырех последовательных событий; (i) приобретение stx2-содержащего бактериофага, (ii) приобретение pO157 и области rfb , (iii) приобретение stx1-содержащего бактериофага и (iv) потеря способности ферментировать D-сорбитол и потеря активности бета-глюкуронидазы (ГУД).
Резервуар для животных
Крупный рогатый скот является основным резервуаром E. coli O157: H7, и этот резервуарный хозяин обычно бессимптомен при переносе этого микроорганизма. У молодых телят этого серотипа наблюдаются редкие случаи диарейных заболеваний. Доля выпадения скота в любой момент времени варьируется. Было обнаружено, что овцы, козы, свиньи и индейки выделяют E. coli O157: H7 с фекалиями.
Молекулярное субтипирование
Для улучшения понимания эпидемиологии E.coli O157: вспышки H7. Эти методы включают гель-электрофорез в импульсном поле (PFGE), полиморфизмы длины рестрикционных фрагментов (RFLP), полиморфизмы длины амплифицированных фрагментов (AFLP) и фаговое типирование [65, 73]. Среди них метод PFGE был стандартизирован CDC и успешно применяется для распознавания связанных со вспышкой, спорадических или неродственных инфекций с 1993 г. [3].
Инфекция
E. coli O157: Инфекция H7 — серьезная проблема общественного здравоохранения в Северной Америке, Европе и других регионах мира.Хотя общее число случаев инфекций E. coli O157: H7 ниже, чем у других кишечных патогенов, таких как Salmonella или Campylobacter spp., Заболевания, вызванные E. coli O157: H7, были намного выше. показатели госпитализации и летальности [43]. Инфекция человека, вызванная E. coli O157: H7, может иметь широкий клинический спектр — от бессимптомных случаев до смерти. Большинство случаев начинаются с диареи без крови и проходят самостоятельно без дальнейших осложнений.Однако у некоторых пациентов кровавая диарея или ГК прогрессирует через 1-3 дня. У 5–10% пациентов с ГК заболевание может прогрессировать до опасных для жизни последствий, ГУС или тромбоцитопенической пурпуры (ТТП) [1]. E. coli O157: H7 — наиболее частая причина ГУС в США. Дети и пожилые люди подвержены повышенному риску серьезных клинических симптомов, таких как ГУС.
Было изучено несколько терапевтических стратегий, включая использование антибиотиков и вакцинацию. Однако не существует специального лечения для E.coli O157: H7 и использование антибиотиков может быть противопоказано. Таким образом, лечение в основном является поддерживающим, чтобы ограничить продолжительность симптомов и предотвратить системные осложнения. Учитывая этот статус, крайне необходимы высокоэффективные меры по профилактике и контролю инфекций E. coli O157: H7 и борьбы с ними.
Передача
В Соединенных Штатах наиболее частым путем передачи инфекции E. coli O157: H7 является через потребление зараженных продуктов питания и воды [56].Однако он также может передаваться напрямую от человека к человеку, особенно в детских дошкольных учреждениях, и от животного к человеку. Были зарегистрированы случаи заражения людей, посещавших контактные зоопарки, молочные фермы или лагеря, где ранее пасся крупный рогатый скот [28, 31]. Недавно сообщалось о потенциальной передаче инфекции воздушным путем в зараженном здании, где находится экспонат с животными [70]. Из 350 вспышек, о которых было сообщено в CDC с 1982 по 2002 годы, пути передачи были определены через пищевые продукты (52%), неизвестные (21%), от человека к человеку (14%), через воду (9%) и при контакте с животными ( 3%) [56].Модель передачи E. coli O157: H7, обновленная по диаграмме Gansheroff и O’Brien [23], показана на. Эти различные пути передачи можно объяснить очень низкой инфекционной дозой (~ 50 КОЕ) E. coli O157: H7.
Передача E. coli O157: H7.
Здоровый крупный рогатый скот является основным резервуаром E. coli O157: H7 и временно переносит этот микроорганизм без симптомов. Загрязненные продукты крупного рогатого скота и зерновые культуры являются преобладающими источниками заражения людей.
Крупный рогатый скот является естественным резервуаром E. coli O157: H7. От 1% до 50% здорового крупного рогатого скота в любой момент времени несут и выделяют E. coli O157: H7 с фекалиями [13, 18, 27]. Загрязненный говяжий фарш является наиболее частым переносчиком вспышек E. coli O157: H7. Продукты из говядины могут быть загрязнены во время убоя, а в процессе измельчения говядины патогенные микроорганизмы могут переноситься с поверхности мяса внутрь. Следовательно, если говяжий фарш приготовлен не полностью, бактерии могут выжить.Кроме того, существует множество зараженных пищевых продуктов, кроме говяжьего фарша, которые были связаны с случаев заражения E. coli O157: H7, включая непастеризованное молоко, питьевую воду, салями, вяленое мясо говядины и свежие продукты, такие как салат, редис. ростки, свежий шпинат и яблочный сидр. Самая крупная вспышка была связана с заражением проростков редиса (1996 г.) в Осаке, Япония, где у 7 966 человек были диагностированы подтвержденные инфекции [45]. Эпидемиологические исследования показывают, что эти пищевые продукты, по-видимому, были заражены фекалиями крупного рогатого скота.Таким образом, предотвращение заражения крупного рогатого скота E. coli O157: H7 может быть одним из наиболее важных методов борьбы. Для борьбы с E. coli O157: H7 на ферме было предложено усовершенствовать практику содержания крупного рогатого скота, выявить ингибирующие корма, иммунизацию, использовать кормовые добавки и использовать пробиотические культуры.
Кислотостойкость
E. coli O157: H7 Кислотостойкость (AR) — это способность бактерий защищаться от чрезвычайно низкого pH ( Низкая инфекционная доза — одна из наиболее известных характеристик E. coli O157: H7, что делает эту бактерию очень заразной.
Низкая инфекционная доза — одна из наиболее известных характеристик E. coli O157: H7, что делает эту бактерию очень заразной.
В различных исследованиях сообщалось об AR штаммов E. coli O157: H7 [5, 12]. Эти исследования определили три эффективных системы AR. Первая система AR требует альтернативного сигма-фактора RpoS и репрессии глюкозы. Мутант rpoS из E. coli O157: H7 выделялся в меньших количествах у экспериментально инфицированных мышей и телят. Вторая система AR требует добавления аргинина во время воздействия кислой среды. Об аргининдекарбоксилазе ( adiA ) и регуляторе adiA ( cysB ) сообщалось во второй системе AR.Третья система AR требует глутамата для защиты в условиях низкого pH. Основные компоненты этой системы AR включают два изофермента глутаматдекарбоксилазы ( gadA или gadB) и предполагаемый антипортер глутамата γ-аминомасляной кислоты ( gadC ). В то время как для защиты требуется только одна из двух глутаматдекарбоксилаз при pH 2,5, оба изофермента глутаматдекарбоксилазы необходимы при pH 2,0. Предыдущие результаты показали, что глутамат-зависимая АР является наиболее эффективной защитой при pH 2.0 в сложной среде. E. coli O157: H7 обладает тремя перекрывающимися системами AR, но контроль и требования к активности AR различны в каждой системе AR.
Помимо этих трех систем AR, было идентифицировано несколько белков, участвующих в AR E. coli O157: H7. Эти белки включают шаперон HdeA, белок, связанный с РНК-полимеразой SspA, и ДНК-связывающий белок Dps. Более того, было показано, что изменения в мембране клеточной стенки или продукции толстой кишки связаны с успешным AR.Таким образом, E. coli O157: H7 использует различные системы AR в зависимости от типа естественной кислой среды.
Колонизация крупного рогатого скота
E. coli O157: H7 естественным образом колонизирует желудочно-кишечный тракт крупного рогатого скота и слизистую оболочку, насыщенную лимфоидными фолликулами, в конечной части прямой кишки, называемой слизистой оболочкой прямой кишки (RAJ), известной как основное место колонизации крупного рогатого скота [39, 48].
Три различных паттерна носительства E. coli O157: H7 у крупного рогатого скота были описаны ранее [14, 39, 58].Во-первых, животные могут быть временно положительными в культуре в течение коротких периодов в несколько дней и считаются пассивными шеддерами и, вероятно, не колонизируются на слизистой оболочке RAJ. Во-вторых, крупный рогатый скот может колонизироваться и выделять бактерии в среднем в течение 1 месяца и обычно не дольше 2 месяцев. В-третьих, некоторые редкие животные колонизируются в течение длительного времени и выделяют бактерии от 3 до 12 месяцев или дольше. Эта уникальная ситуация, при которой у нескольких животных развивается длительная колонизация (> 2 месяцев) E.coli O157: H7, вероятно, из-за бактериальной ассоциации на слизистой оболочке RAJ; однако это может быть связано с уникальной колонизацией бактериями определенного участка (а) в дополнение к слизистой оболочке RAJ.
Возраст, диета и иммунитет отдельных особей крупного рогатого скота также потенциально могут влиять на колонизацию бактерий. Cray and Moon [14] сообщили, что телята выделяют E. coli O157: H7 дольше, чем взрослый крупный рогатый скот при таком же уровне инокулята E. coli O157: H7.
Снижение уровня перевозки E.coli O157: H7 у крупного рогатого скота, как основной источник инфекции E. coli O157: H7, будет играть ключевую роль в снижении риска заражения человека. Понимание факторов колонизации E. coli O157: H7 будет необходимо для разработки эффективных стратегий снижения или предотвращения носительства E. coli O157: H7 коровами.
Экологическое выживание
E. coli O157: H7 может выживать и сохраняться в различных средах, таких как почва, вода и еда, а также в резервуарах для животных (). E. coli O157: H7, как было показано, выживает в течение года в почве, обработанной навозом, и в течение 21 месяца в необработанном навозе, который не подвергался компостированию [30]. Компостирование навоза эффективно при уничтожении E. coli O157: H7, если температура поддерживается выше 50 ° C в течение 6 дней. E. coli O157: H7 может долгое время выживать в воде, особенно при низких температурах. Отложения в водоемах, загрязненные фекалиями крупного рогатого скота, могут служить долгосрочным (> 8 месяцев) резервуаром E.coli O157: H7, а выжившие бактерии в загрязненных желобах являются источником инфекции [38]. Баркер и др. . [2] показали, что E. coli O157: H7 выживает и реплицируется в Acanthamoeba polyphaga. A. polyphaga — обычное простейшее в окружающей среде, которое широко распространено в почве, воде и фекалиях. Таким образом, он может быть эффективным средством передачи E. coli O157: H7 в этих средах.
Экологическая схема E.coli O157: H7.
ЖКТ, желудочно-кишечный тракт.
Чтобы выжить в различных средах, E. coli O157: H7 требуется способность адаптироваться к колебаниям или резким изменениям температуры, pH и осмолярности, которые обычно встречаются в природе. Например, продукция экзополисахарида (EPS) E. coli O157: H7 связана с устойчивостью к теплу и кислоте, а изменение липидного состава мембран индуцируется тепловым стрессом [77].
Эти экологические адаптации E. coli O157: H7 играют важную роль в сохранении и распространении этого микроорганизма на фермах и в увеличивающейся передаче от крупного рогатого скота к крупному рогатому скоту. Кроме того, способность выживать за пределами резервуара-хозяина увеличивает риск того, что патоген может заразить посевы и произвести через заражение коровьим навозом, орошение зараженной водой или прямой контакт с инфицированными животными [42].
Основные факторы вирулентности
Определение факторов и механизмов вирулентности E.coli O157: Патогенез H7 был предметом многочисленных исследований (). Производство Stx считается важным, но не единственно ответственным за заболевание. Кроме того, E. coli O157: H7, ассоциированный с тяжелым заболеванием человека, должен колонизировать слизистую кишечника, а наличие pO157 также коррелирует со способностью вызывать заболевание. Каждый из этих аспектов описан ниже.
Факторы вирулентности E. coli O157: H7.
Shiga Toxins (Stxs)
Stx — мощный цитотоксин, кодируемый бактериофагом.Stx расширяется из единственной транскрипционной единицы и вызывает повреждение множества типов клеток [29]. Stx можно разделить на две группы, называемые Stx1 и Stx2, но они не генерируют перекрестно-реактивные антитела, гомологичные по аминокислотным последовательностям на 56%. Stx1 идентичен Stx из Shigella dysenteriae I, но с одной аминокислотной разницей. Вирулентные изоляты E. coli O157: H7 могут экспрессировать только Stx1, только Stx2 или оба токсина. Известно, что Stx2 более токсичен и чаще ассоциируется с HC или HUS при инфекциях человека, чем штаммы Stx1 [6, 50].
Stx имеет консервативную структуру, состоящую из одной ферментативно активной субъединицы А (А1) и пяти идентичных рецептор-связывающих субъединиц В (В5). Субъединица B5 связывается со специфическими рецепторами хозяина глоботриаозилцерамидом (Gb3) или глоботетраозилцерамидом (Gb4) [47]. После связывания Stx (A1B5) с клеткой-хозяином субъединица A интернализуется в цитоплазму. Al ингибирует синтез белка путем специфического удаления одного остатка аденина из 28S рРНК 60S субъединицы рибосомы [59]. Подробные механизмы транслокации Stx в различные ткани до конца не изучены.
Локус поражения энтероцитов
E. coli O157: H7 колонизирует слизистую оболочку кишечника и вызывает характерное гистопатологическое поражение, известное как прикрепляющиеся и размывающие (A / E) поражения. Поражение A / E характеризуется стиранием микроворсинок и прикреплением бактерий к мембране эпителиальной клетки. Присоединенные бактерии стимулируют накопление актина в клетках-хозяевах, что приводит к возвышению пьедестала прикрепления [11]. Генетические исследования показали, что гены, ответственные за поражения A / E, отображаются в 13-м регионе, который был обозначен как локус сглаживания энтероцитов (LEE).LEE E. coli O157: H7 также консервативен в EPEC, и хорошо известно, что присутствие LEE тесно связано с заболеванием [24]. LEE E. coli O157: H7 имеет размер 43 т.п.н. и содержит дополнительную последовательность профага в 7,5 т.п.н. по сравнению со штаммами ЕРЕС. Роль этой дополнительной последовательности четко не определена. LEE состоит по крайней мере из 41 различных генов, организованных в три основных региона; (i) система секреции типа III (TTSS), которая экспортирует эффективные молекулы; (ii) адгезия под названием интимин и его транслоцированный рецептор Tir, который транслоцируется в мембрану клетки-хозяина с помощью TTSS; и (iii) несколько секретируемых белков (Esp) как часть TTSS, которые важны для модификации передачи сигнала клетки-хозяина во время образования повреждений A / E [15, 52].Недавно были идентифицированы эффекторы, не кодируемые LEE, и выяснение их роли еще больше расширит понимание патологических явлений при инфекциях E. coli O157: H7 [16].
Плазмида O157 (pO157)
В дополнение к Stxs и LEE, которые оба хромосомно кодируются, все клинические изоляты E. coli O157: H7 обладают предполагаемой плазмидой вирулентности, называемой pO157.
Плазмида O157 (pO157)
Плазмиды
Плазмида представляет собой внехромосомную ДНК, способную к репликации независимо от хромосомной ДНК.Плазмиды — это подвижные элементы, которые обеспечивают различные полезные свойства хозяина, такие как устойчивость к антибиотикам и тяжелым металлам, выработка токсинов и других факторов вирулентности, биотрансформация углеводородов и симбиотическая азотфиксация [22]. Кодируемые плазмидой гены необходимы для полного патогенеза многих энтеропатогенных бактерий, включая виды Shigella , Yersinia , Salmonella и E. coli .
pO157
E.coli O157: H7 содержит высококонсервативную плазмиду, названную pO157. PO157 представляет собой неконъюгативную F-подобную плазмиду с размером диапазона от 92 до 104 т.п.н. Полная последовательность pO157 в двух различных изолятах вспышки была опубликована [10, 41]. PO157 показывает динамическую структуру и включает в себя различные мобильные генетические элементы, такие как транспозоны, профаги, инсерционные последовательности (IS) и части других плазмид. Гетерогенный состав pO157 может ограничивать совместные ответы на функциональные области pO157.Среди них IS или остатки IS часто связаны с сегментами, связанными с вирулентностью, которые аналогичны составам большой плазмиды вирулентности в Shigella spp. [10, 41]. Эти результаты показывают, что фактический pO157 образуется в результате интеграции фрагментов из эволюционно различных видов в F-подобную плазмиду, и, таким образом, факторы вирулентности или предполагаемые факторы вирулентности на различных сегментах pO157 могут иметь различное происхождение. Полная последовательность pO157 обнаруживает 100 открытых рамок считывания (ORF) [10].Среди них 43 ORF показали достаточное сходство с известными белками, предполагая функции, а 22 ORF не имели убедительного сходства с какими-либо известными белками. Тридцать пять белков предположительно участвуют в патогенезе инфекций E. coli O157: H7, но из них только 19 генов были ранее охарактеризованы, включая гемолизин ( ehxA ) [63], каталазу-пероксидазу ( katP ) [9], устройство системы секреции типа II ( etp ) [62], сериновая протеаза ( espP ) [8], предполагаемый адгезин ( toxB ) [67], металлопротеиназа цинка (s tcE ) [37] и консервативный фрагмент eae ( ecf ) [75].Однако биологическое значение pO157 в патогенезе до конца не изучено.
Гемолизин (
ehx )
 Другие статьи в PMC, в которых цитируется опубликованная статья.
Другие статьи в PMC, в которых цитируется опубликованная статья. В этом обзоре представлен обзор платы E. coli O157: H7 с акцентом на pO157.
В этом обзоре представлен обзор платы E. coli O157: H7 с акцентом на pO157. coli , которые продуцируют токсины Шига (Stxs) и вызывают геморрагический колит (HC) и опасные для жизни последствия гемолитико-уремического синдрома (HUS) у людей. . Некоторые серотипы в EHEC часто связаны с заболеваниями человека, такими как O26: h21, O91: h31, O111: H8, O157: NM и O157: H7 [44, 51]. E. coli O157: H7 — серотип EHEC, наиболее часто выделяемый у больных в США, Японии и Великобритании, и он является предметом настоящего обзора.
coli , которые продуцируют токсины Шига (Stxs) и вызывают геморрагический колит (HC) и опасные для жизни последствия гемолитико-уремического синдрома (HUS) у людей. . Некоторые серотипы в EHEC часто связаны с заболеваниями человека, такими как O26: h21, O91: h31, O111: H8, O157: NM и O157: H7 [44, 51]. E. coli O157: H7 — серотип EHEC, наиболее часто выделяемый у больных в США, Японии и Великобритании, и он является предметом настоящего обзора. coli O157: инфекции H7 вызывают 73 000 заболеваний, 2200 госпитализаций и 60 смертей ежегодно в США [43]. Данные эпиднадзора за вспышками, предоставленные CDC, показали, что инфекций E. coli O157: H7 сокращаются после пика в 1999 г. Однако крупные вспышки и спорадические случаи продолжают иметь место. Ежегодные затраты на заболевание, вызванное инфекциями E. coli O157: H7, составили 405 миллионов долларов, включая потерю производительности, медицинское обслуживание и преждевременную смерть [21]. Высокая стоимость болезни требует дополнительных усилий по борьбе с этим возбудителем.
coli O157: инфекции H7 вызывают 73 000 заболеваний, 2200 госпитализаций и 60 смертей ежегодно в США [43]. Данные эпиднадзора за вспышками, предоставленные CDC, показали, что инфекций E. coli O157: H7 сокращаются после пика в 1999 г. Однако крупные вспышки и спорадические случаи продолжают иметь место. Ежегодные затраты на заболевание, вызванное инфекциями E. coli O157: H7, составили 405 миллионов долларов, включая потерю производительности, медицинское обслуживание и преждевременную смерть [21]. Высокая стоимость болезни требует дополнительных усилий по борьбе с этим возбудителем. Таким образом, агар Sorbitol MacConkey (SMAC) с добавлением MUG был использован для обнаружения E. coli O157: H7. Для увеличения селективности на E.coli O157: H7, цефиксим, теллурит калия и ванкомицин были добавлены в чашки с агаром SMAC для подавления другой грамотрицательной флоры. Серотипы O157 и H7 могут быть дополнительно подтверждены коммерчески доступным анализом латексной агглютинации.
Таким образом, агар Sorbitol MacConkey (SMAC) с добавлением MUG был использован для обнаружения E. coli O157: H7. Для увеличения селективности на E.coli O157: H7, цефиксим, теллурит калия и ванкомицин были добавлены в чашки с агаром SMAC для подавления другой грамотрицательной флоры. Серотипы O157 и H7 могут быть дополнительно подтверждены коммерчески доступным анализом латексной агглютинации. coli O157: H7 (1,4 МБ) представляют собой горизонтально переносимые чужеродные ДНК, такие как профаг и профагоподобные элементы. E. coli O157: H7 содержит 463 фаго-ассоциированных гена по сравнению с 29 в E.coli K-12 [72]. Изменение содержания G + C является одним из признаков того, что геномная область была получена путем горизонтального переноса, и Putonti et al. [55] подсчитал, что по крайней мере 53 различных вида внесли вклад в эти уникальные последовательности в E. coli O157: H7. Связанные с вирулентностью гены между двумя секвенированными штаммами E. coli O157: H7 почти идентичны (99%). Очевидно, что как приобретение, так и потеря ДНК сыграли важную роль в эволюции патогенеза E.coli O157: H7.
coli O157: H7 (1,4 МБ) представляют собой горизонтально переносимые чужеродные ДНК, такие как профаг и профагоподобные элементы. E. coli O157: H7 содержит 463 фаго-ассоциированных гена по сравнению с 29 в E.coli K-12 [72]. Изменение содержания G + C является одним из признаков того, что геномная область была получена путем горизонтального переноса, и Putonti et al. [55] подсчитал, что по крайней мере 53 различных вида внесли вклад в эти уникальные последовательности в E. coli O157: H7. Связанные с вирулентностью гены между двумя секвенированными штаммами E. coli O157: H7 почти идентичны (99%). Очевидно, что как приобретение, так и потеря ДНК сыграли важную роль в эволюции патогенеза E.coli O157: H7. E. coli O15: H7 возник в результате четырех последовательных событий; (i) приобретение stx2-содержащего бактериофага, (ii) приобретение pO157 и области rfb , (iii) приобретение stx1-содержащего бактериофага и (iv) потеря способности ферментировать D-сорбитол и потеря активности бета-глюкуронидазы (ГУД).
E. coli O15: H7 возник в результате четырех последовательных событий; (i) приобретение stx2-содержащего бактериофага, (ii) приобретение pO157 и области rfb , (iii) приобретение stx1-содержащего бактериофага и (iv) потеря способности ферментировать D-сорбитол и потеря активности бета-глюкуронидазы (ГУД). Среди них метод PFGE был стандартизирован CDC и успешно применяется для распознавания связанных со вспышкой, спорадических или неродственных инфекций с 1993 г. [3].
Среди них метод PFGE был стандартизирован CDC и успешно применяется для распознавания связанных со вспышкой, спорадических или неродственных инфекций с 1993 г. [3]. E. coli O157: H7 — наиболее частая причина ГУС в США. Дети и пожилые люди подвержены повышенному риску серьезных клинических симптомов, таких как ГУС.
E. coli O157: H7 — наиболее частая причина ГУС в США. Дети и пожилые люди подвержены повышенному риску серьезных клинических симптомов, таких как ГУС. Были зарегистрированы случаи заражения людей, посещавших контактные зоопарки, молочные фермы или лагеря, где ранее пасся крупный рогатый скот [28, 31]. Недавно сообщалось о потенциальной передаче инфекции воздушным путем в зараженном здании, где находится экспонат с животными [70]. Из 350 вспышек, о которых было сообщено в CDC с 1982 по 2002 годы, пути передачи были определены через пищевые продукты (52%), неизвестные (21%), от человека к человеку (14%), через воду (9%) и при контакте с животными ( 3%) [56].Модель передачи E. coli O157: H7, обновленная по диаграмме Gansheroff и O’Brien [23], показана на. Эти различные пути передачи можно объяснить очень низкой инфекционной дозой (~ 50 КОЕ) E. coli O157: H7.
Были зарегистрированы случаи заражения людей, посещавших контактные зоопарки, молочные фермы или лагеря, где ранее пасся крупный рогатый скот [28, 31]. Недавно сообщалось о потенциальной передаче инфекции воздушным путем в зараженном здании, где находится экспонат с животными [70]. Из 350 вспышек, о которых было сообщено в CDC с 1982 по 2002 годы, пути передачи были определены через пищевые продукты (52%), неизвестные (21%), от человека к человеку (14%), через воду (9%) и при контакте с животными ( 3%) [56].Модель передачи E. coli O157: H7, обновленная по диаграмме Gansheroff и O’Brien [23], показана на. Эти различные пути передачи можно объяснить очень низкой инфекционной дозой (~ 50 КОЕ) E. coli O157: H7.
 Эпидемиологические исследования показывают, что эти пищевые продукты, по-видимому, были заражены фекалиями крупного рогатого скота.Таким образом, предотвращение заражения крупного рогатого скота E. coli O157: H7 может быть одним из наиболее важных методов борьбы. Для борьбы с E. coli O157: H7 на ферме было предложено усовершенствовать практику содержания крупного рогатого скота, выявить ингибирующие корма, иммунизацию, использовать кормовые добавки и использовать пробиотические культуры. Низкая инфекционная доза — одна из наиболее известных характеристик E. coli O157: H7, что делает эту бактерию очень заразной.
Эпидемиологические исследования показывают, что эти пищевые продукты, по-видимому, были заражены фекалиями крупного рогатого скота.Таким образом, предотвращение заражения крупного рогатого скота E. coli O157: H7 может быть одним из наиболее важных методов борьбы. Для борьбы с E. coli O157: H7 на ферме было предложено усовершенствовать практику содержания крупного рогатого скота, выявить ингибирующие корма, иммунизацию, использовать кормовые добавки и использовать пробиотические культуры. Низкая инфекционная доза — одна из наиболее известных характеристик E. coli O157: H7, что делает эту бактерию очень заразной. Предыдущие результаты показали, что глутамат-зависимая АР является наиболее эффективной защитой при pH 2.0 в сложной среде. E. coli O157: H7 обладает тремя перекрывающимися системами AR, но контроль и требования к активности AR различны в каждой системе AR.
Предыдущие результаты показали, что глутамат-зависимая АР является наиболее эффективной защитой при pH 2.0 в сложной среде. E. coli O157: H7 обладает тремя перекрывающимися системами AR, но контроль и требования к активности AR различны в каждой системе AR.
 Cray and Moon [14] сообщили, что телята выделяют E. coli O157: H7 дольше, чем взрослый крупный рогатый скот при таком же уровне инокулята E. coli O157: H7.
Cray and Moon [14] сообщили, что телята выделяют E. coli O157: H7 дольше, чем взрослый крупный рогатый скот при таком же уровне инокулята E. coli O157: H7. coli O157: H7, если температура поддерживается выше 50 ° C в течение 6 дней. E. coli O157: H7 может долгое время выживать в воде, особенно при низких температурах. Отложения в водоемах, загрязненные фекалиями крупного рогатого скота, могут служить долгосрочным (> 8 месяцев) резервуаром E.coli O157: H7, а выжившие бактерии в загрязненных желобах являются источником инфекции [38]. Баркер и др. . [2] показали, что E. coli O157: H7 выживает и реплицируется в Acanthamoeba polyphaga. A. polyphaga — обычное простейшее в окружающей среде, которое широко распространено в почве, воде и фекалиях. Таким образом, он может быть эффективным средством передачи E. coli O157: H7 в этих средах.
coli O157: H7, если температура поддерживается выше 50 ° C в течение 6 дней. E. coli O157: H7 может долгое время выживать в воде, особенно при низких температурах. Отложения в водоемах, загрязненные фекалиями крупного рогатого скота, могут служить долгосрочным (> 8 месяцев) резервуаром E.coli O157: H7, а выжившие бактерии в загрязненных желобах являются источником инфекции [38]. Баркер и др. . [2] показали, что E. coli O157: H7 выживает и реплицируется в Acanthamoeba polyphaga. A. polyphaga — обычное простейшее в окружающей среде, которое широко распространено в почве, воде и фекалиях. Таким образом, он может быть эффективным средством передачи E. coli O157: H7 в этих средах. Например, продукция экзополисахарида (EPS) E. coli O157: H7 связана с устойчивостью к теплу и кислоте, а изменение липидного состава мембран индуцируется тепловым стрессом [77].
Например, продукция экзополисахарида (EPS) E. coli O157: H7 связана с устойчивостью к теплу и кислоте, а изменение липидного состава мембран индуцируется тепловым стрессом [77]. Каждый из этих аспектов описан ниже.
Каждый из этих аспектов описан ниже. После связывания Stx (A1B5) с клеткой-хозяином субъединица A интернализуется в цитоплазму. Al ингибирует синтез белка путем специфического удаления одного остатка аденина из 28S рРНК 60S субъединицы рибосомы [59]. Подробные механизмы транслокации Stx в различные ткани до конца не изучены.
После связывания Stx (A1B5) с клеткой-хозяином субъединица A интернализуется в цитоплазму. Al ингибирует синтез белка путем специфического удаления одного остатка аденина из 28S рРНК 60S субъединицы рибосомы [59]. Подробные механизмы транслокации Stx в различные ткани до конца не изучены. LEE E. coli O157: H7 имеет размер 43 т.п.н. и содержит дополнительную последовательность профага в 7,5 т.п.н. по сравнению со штаммами ЕРЕС. Роль этой дополнительной последовательности четко не определена. LEE состоит по крайней мере из 41 различных генов, организованных в три основных региона; (i) система секреции типа III (TTSS), которая экспортирует эффективные молекулы; (ii) адгезия под названием интимин и его транслоцированный рецептор Tir, который транслоцируется в мембрану клетки-хозяина с помощью TTSS; и (iii) несколько секретируемых белков (Esp) как часть TTSS, которые важны для модификации передачи сигнала клетки-хозяина во время образования повреждений A / E [15, 52].Недавно были идентифицированы эффекторы, не кодируемые LEE, и выяснение их роли еще больше расширит понимание патологических явлений при инфекциях E. coli O157: H7 [16].
LEE E. coli O157: H7 имеет размер 43 т.п.н. и содержит дополнительную последовательность профага в 7,5 т.п.н. по сравнению со штаммами ЕРЕС. Роль этой дополнительной последовательности четко не определена. LEE состоит по крайней мере из 41 различных генов, организованных в три основных региона; (i) система секреции типа III (TTSS), которая экспортирует эффективные молекулы; (ii) адгезия под названием интимин и его транслоцированный рецептор Tir, который транслоцируется в мембрану клетки-хозяина с помощью TTSS; и (iii) несколько секретируемых белков (Esp) как часть TTSS, которые важны для модификации передачи сигнала клетки-хозяина во время образования повреждений A / E [15, 52].Недавно были идентифицированы эффекторы, не кодируемые LEE, и выяснение их роли еще больше расширит понимание патологических явлений при инфекциях E. coli O157: H7 [16]. coli O157: H7 обладают предполагаемой плазмидой вирулентности, называемой pO157.
coli O157: H7 обладают предполагаемой плазмидой вирулентности, называемой pO157. PO157 показывает динамическую структуру и включает в себя различные мобильные генетические элементы, такие как транспозоны, профаги, инсерционные последовательности (IS) и части других плазмид. Гетерогенный состав pO157 может ограничивать совместные ответы на функциональные области pO157.Среди них IS или остатки IS часто связаны с сегментами, связанными с вирулентностью, которые аналогичны составам большой плазмиды вирулентности в Shigella spp. [10, 41]. Эти результаты показывают, что фактический pO157 образуется в результате интеграции фрагментов из эволюционно различных видов в F-подобную плазмиду, и, таким образом, факторы вирулентности или предполагаемые факторы вирулентности на различных сегментах pO157 могут иметь различное происхождение. Полная последовательность pO157 обнаруживает 100 открытых рамок считывания (ORF) [10].Среди них 43 ORF показали достаточное сходство с известными белками, предполагая функции, а 22 ORF не имели убедительного сходства с какими-либо известными белками.
PO157 показывает динамическую структуру и включает в себя различные мобильные генетические элементы, такие как транспозоны, профаги, инсерционные последовательности (IS) и части других плазмид. Гетерогенный состав pO157 может ограничивать совместные ответы на функциональные области pO157.Среди них IS или остатки IS часто связаны с сегментами, связанными с вирулентностью, которые аналогичны составам большой плазмиды вирулентности в Shigella spp. [10, 41]. Эти результаты показывают, что фактический pO157 образуется в результате интеграции фрагментов из эволюционно различных видов в F-подобную плазмиду, и, таким образом, факторы вирулентности или предполагаемые факторы вирулентности на различных сегментах pO157 могут иметь различное происхождение. Полная последовательность pO157 обнаруживает 100 открытых рамок считывания (ORF) [10].Среди них 43 ORF показали достаточное сходство с известными белками, предполагая функции, а 22 ORF не имели убедительного сходства с какими-либо известными белками. Тридцать пять белков предположительно участвуют в патогенезе инфекций E. coli O157: H7, но из них только 19 генов были ранее охарактеризованы, включая гемолизин ( ehxA ) [63], каталазу-пероксидазу ( katP ) [9], устройство системы секреции типа II ( etp ) [62], сериновая протеаза ( espP ) [8], предполагаемый адгезин ( toxB ) [67], металлопротеиназа цинка (s tcE ) [37] и консервативный фрагмент eae ( ecf ) [75].Однако биологическое значение pO157 в патогенезе до конца не изучено.
Тридцать пять белков предположительно участвуют в патогенезе инфекций E. coli O157: H7, но из них только 19 генов были ранее охарактеризованы, включая гемолизин ( ehxA ) [63], каталазу-пероксидазу ( katP ) [9], устройство системы секреции типа II ( etp ) [62], сериновая протеаза ( espP ) [8], предполагаемый адгезин ( toxB ) [67], металлопротеиназа цинка (s tcE ) [37] и консервативный фрагмент eae ( ecf ) [75].Однако биологическое значение pO157 в патогенезе до конца не изучено. Гемолизин был первым описанным фактором вирулентности pO157 [4, 61]. Оперон гемолизина ( ehxCABD ) может быть чужеродным по происхождению, потому что он имеет другой процент G + C и использование кодонов, чем окружающее генетическое содержимое. Фрагмент размером 3,4 т.п.н. кодирует гены, необходимые для синтеза и транспорта гемолизина, и эта область была использована в качестве диагностического зонда для E. coli O157: H7 и часто изоляты EHEC. Несколько исследований показали, что гемолизин высоко консервативен среди различных серотипов EHEC, таких как O157: H7, O111: H8 и O8: h29, но не известно, имеют ли они идентичные биологические активности [7].
coli O157: H7 и часто изоляты EHEC. Несколько исследований показали, что гемолизин высоко консервативен среди различных серотипов EHEC, таких как O157: H7, O111: H8 и O8: h29, но не известно, имеют ли они идентичные биологические активности [7].
Каталаза-пероксидаза (
katP ) Ген активности каталазы-пероксидазы ( katP ) был идентифицирован из pO157 [9]. Этот ген имеет размер 2,2 т.п.н. и очень гомологичен бактериальной бифункциональной каталазе-пероксидазе.Активность фермента KatP E. coli O157: H7 была показана как во фракциях цитоплазмы, так и во фракциях периплазмы. N-концевая сигнальная последовательность предполагает, что этот фермент транспортируется через цитоплазматическую мембрану. Ген katP был обнаружен во всех штаммах E. coli O157: H7, но не обнаружен в штаммах EPEC, ETEC, EIEC и EAggEC. Этот фермент может помочь E. coli O157: H7 колонизировать кишечник хозяина за счет снижения окислительного стресса и использования побочного продукта кислорода в условиях пониженного или дефицитного кислорода в кишечнике хозяина.
Система секреции типа II (T2SS) (
etp )pO157 кодирует 13 открытых рамок считывания с именами от etpC до etpO , которые демонстрируют большое сходство с T2SS грамотрицательных бактерий [62]. Эти гены расположены рядом с локусом гемолизина. Инсерционный элемент, подобный IS911, был обнаружен далеко от генов etp и ehx . Подобно гену katP , гены etp были также обнаружены во всех штаммах E. coli O157: H7, некоторые — в штаммах EHEC, отличных от O157, и не обнаружены в штаммах EPEC, ETEC, EIEC и EAggEC.Этот T2SS аналогичен пути секреции пуллуланазы ( pulO ) Klebsiella oxytoca , но его функция не идентифицирована.
Сериновая протеаза (
espP ) EspP представляет собой кодируемую pO157 сериновую протеазу типа V, которая, как известно, расщепляет пепсин A и фактор свертывания крови V человека [8]. Этот внеклеточный фермент подобен нескольким секретируемым или поверхностно связанным белкам, включая PssA в EHEC O26: H-, EspC в EPEC и протеазу IgA1 у видов Neisseria [69]. Недавно Dziva et al . [19] сообщили, что EspP влияет на колонизацию кишечника телят и прилипание к первичным кишечным эпителиальным клеткам крупного рогатого скота. Более того, деградация человеческого фактора свертывания V через EspP может способствовать кровотечению из слизистой оболочки, наблюдаемому у пациентов с HC.
Недавно Dziva et al . [19] сообщили, что EspP влияет на колонизацию кишечника телят и прилипание к первичным кишечным эпителиальным клеткам крупного рогатого скота. Более того, деградация человеческого фактора свертывания V через EspP может способствовать кровотечению из слизистой оболочки, наблюдаемому у пациентов с HC.
Металлопротеаза (
stcE ) Металлопротеиназа, StcE, кодируется на pO157 и специфически расщепляет ингибитор C1-эстеразы [37]. Ингибитор С1-эстеразы является регулятором множества протеолитических каскадов, связанных с путями воспаления, такими как классический комплемент, внутренняя коагуляция и контактная активация.StcE секретируется через T2SS, кодируемый на pO157, и регулируется регулятором, кодируемым LEE ( ler ) [20, 37]. Grys и др. . [25] продемонстрировали, что StcE может способствовать интимной адгезии E. coli O157: H7 к клеткам Hep2 in vitro . Ген stcE был обнаружен все в E. coli O157: H7, некоторые в серотипе EPEC O55: H7, и он не обнаружен в других диареи E. coli .
coli O157: H7, некоторые в серотипе EPEC O55: H7, и он не обнаружен в других диареи E. coli .
Предполагаемая адгезия (
toxB ) Ген toxB кодируется в последовательности 9.Размером 5 т.п.н., и его предсказанный продукт показывает 20% сходство с токсином B Clostridium difficile [41]. Недавние исследования показали, что ToxB способствует прикреплению E. coli O157: H7 к клеткам Caco-2 за счет повышенной секреции TTSS [67]. Более того, сравнение последовательностей показало, что ToxB имеет 28% идентичности аминокислот и 47% сходства с предсказанным продуктом efa-1 / lifA , другого гена вирулентности, часто обнаруживаемого на хромосоме EPEC и не-O157 EHEC. изоляты [46].Присутствие гена efa-1 / lifA , как известно, ингибирует активацию желудочно-кишечных лимфоцитов человека и мыши, и, следовательно, ToxB может участвовать в ингибировании лимфоцитов хозяина [36]. Однако мутация генов toxB и efa-1 не влияла на колонизацию кишечника у телят или овец [66].
Ген-положительные консервативные фрагменты Eae (
ecf ) Недавно мы сообщили, что pO157 кодирует оперон ecf ( ecf1 — 4 ), температура которого регулируется внутренне изогнутой ДНК [76]. ecf1 и ecf2 кодируют предполагаемую полисахариддеацетилазу и ЛПС α-1, 7- N -ацетилглюкозаминтрансферазу, соответственно, и оба являются уникальными для pO157 [32]. ecf3 демонстрирует сходство с предполагаемым белком внешней мембраны в E. coli K1, ассоциированным с бактериальной инвазией [49]. ecf4 , также называемый msbB2 , кодирует вторую копию липид А миристоилтрансферазы [35, 76]. Двойной мутант, несущий делеции в ecf4 и его хромосомную копию lpxM из E.coli O157: H7 имеет измененную структуру липида А и состав мембранных жирных кислот, а также демонстрирует пониженную стойкость в желудочно-кишечном тракте крупного рогатого скота [76]. Однако единственный мутант ecf4 не показал значительных различий по сравнению с E. coli дикого типа O157: H7.
coli дикого типа O157: H7.
Патогенез pO157
После первого сообщения о том, что pO157 необходим для экспрессии фимбрий и адгезии к эпителиальным клеткам, несколько исследований сообщили о противоречивых результатах о роли pO157 в прикреплении к эпителиальным клеткам () [74]. Исследования pO157 in vivo на животных моделях, включая мышей, кроликов и гнобиотических поросят, также показали противоречивые результаты. Однако исследования in vivo имеют ограничения, поскольку не существует подходящей модели на животных, воспроизводящей все аспекты заболевания. Следовательно, точную роль pO157 в патогенезе E. coli O157: H7 еще предстоит определить.
Таблица 1
Резюме исследований патогенеза pO157 in vitro и in vivo .
| Год | Мишень | Патогенез | Эффект | ||||
|---|---|---|---|---|---|---|---|
| In vitro | 1987 | Целая плазмида41 | эпитрииЦелая плазмида | эпитерия | Экспрессия эпителиальных клеток 909|||
| 1990 | Целая плазмида | Адгезия к эпителиальным клеткам | Да | ||||
| 1993 | Вся плазмида | Производство пилли | Нет | Адгезия к эпителиальным клеткам | Да | ||
| 2005 | Ген stcE на pO157 | Прилипание к эпителиальным клеткам | 909P29 909P 29 esp, 2007 | Приверженность t o первичные кишечные эпителиальные клетки крупного рогатого скота | Да | ||
| In vivo | 1987 | Целая плазмида | Прикрепление и удаление очагов поражения у гнотобиотических поросят | Нет | 1990 | 909 Плазмида мышиНет | |
| 1993 | Целая плазмида | Клинические симптомы у кролика | Нет | ||||
| 2006 | Вся плазмида | Колонизация крупного рогатого скота | Да | Whole крупного рогатого скота | Да | ||
| 2007 | Ген espP на pO157 | Колонизация телят | Да |
Недавно мы показали, что pO157 влияет на эффективность E. coli O157: H7 колонизация здорового скота и выживание в кислых условиях [40, 64]. Изогенный мутант E. coli O157: H7 ΔpO157 более устойчив к кислой синтетической желудочной жидкости крупного рогатого скота и желчи, чем дикий тип [40]. Эта повышенная кислотная устойчивость мутанта ΔpO157 обусловлена повышенной экспрессией глутаматдекарбоксилазы (GAD). Механизм регуляции gad с помощью pO157 неизвестен, но, вероятно, связан с регуляцией pO157 хромосомных генов. In vivo , мутант ΔpO157 выживает при прохождении через желудочно-кишечный тракт крупного рогатого скота лучше, чем дикий тип, но не колонизирует слизистую оболочку RAJ крупного рогатого скота так же, как дикий тип [40, 64].
coli O157: H7 колонизация здорового скота и выживание в кислых условиях [40, 64]. Изогенный мутант E. coli O157: H7 ΔpO157 более устойчив к кислой синтетической желудочной жидкости крупного рогатого скота и желчи, чем дикий тип [40]. Эта повышенная кислотная устойчивость мутанта ΔpO157 обусловлена повышенной экспрессией глутаматдекарбоксилазы (GAD). Механизм регуляции gad с помощью pO157 неизвестен, но, вероятно, связан с регуляцией pO157 хромосомных генов. In vivo , мутант ΔpO157 выживает при прохождении через желудочно-кишечный тракт крупного рогатого скота лучше, чем дикий тип, но не колонизирует слизистую оболочку RAJ крупного рогатого скота так же, как дикий тип [40, 64].
pO157-подобные плазмиды в EHEC
Большие плазмиды, похожие на pO157, обнаружены в большинстве изолятов EHEC, отличных от O157, но не во всех изолятах от человека, и размер варьируется от 70 до 200 т.п.н. [26]. Эти плазмиды обычно несут оперон гемолизина ( ehx ), но гены etpC-O , katP и espP встречаются менее чем в 50% изолятов [11]. Сообщается, что некоторые из этих плазмид EHEC-гемолизин участвуют в адгезии, но некоторые нет.Эпидемиологические данные свидетельствуют о более сильной корреляции присутствия этой плазмиды EHEC-гемолизин с развитием ГУС, а не с диареей. Помимо плазмид pO157 или EHEC-гемолизин, в изолятах E. coli O157: H7 был обнаружен ряд других плазмид размером от 2 до 87 т.п.н. Однако не было обнаружено корреляции с наличием какой-либо из этих плазмид и клиническим заболеванием.
Сообщается, что некоторые из этих плазмид EHEC-гемолизин участвуют в адгезии, но некоторые нет.Эпидемиологические данные свидетельствуют о более сильной корреляции присутствия этой плазмиды EHEC-гемолизин с развитием ГУС, а не с диареей. Помимо плазмид pO157 или EHEC-гемолизин, в изолятах E. coli O157: H7 был обнаружен ряд других плазмид размером от 2 до 87 т.п.н. Однако не было обнаружено корреляции с наличием какой-либо из этих плазмид и клиническим заболеванием.
Заключительные замечания
Этот обзор посвящен серотипу E.coli O157: H7 и его плазмида размером 92 т.п.н. E. coli O157: H7 вызывает тяжелые заболевания людей во всем мире. Три основных фактора вирулентности включают токсины Шига, продукты острова патогенности, называемые локусом сглаживания энтероцитов, и продукты F-подобной плазмиды pO157. Этот патоген хорошо выживает в различных средах, от своего бесшумного резервуара в здоровом скоте до окружающей среды на ферме. Гены, кодируемые pO157, влияют на прикрепление бактерий к эукариотическим клеткам, колонизацию крупного рогатого скота и устойчивость к кислотам. Дальнейшие исследования для понимания механизмов патогенеза и устойчивости E. coli O157: H7 в окружающей среде позволят разработать эффективные меры по предотвращению заболеваний человека.
Дальнейшие исследования для понимания механизмов патогенеза и устойчивости E. coli O157: H7 в окружающей среде позволят разработать эффективные меры по предотвращению заболеваний человека.
Ссылки
1. Банатвала Н., Гриффин П.М., Грин К.Д., Барретт Т.Дж., Бибб В.Ф., Грин Дж. Х., Уэллс Дж. Дж. Национальное проспективное исследование гемолитико-уремического синдрома США: микробиологические, серологические, клинические и эпидемиологические данные. J Infect Dis. 2001; 183: 1063–1070. [PubMed] [Google Scholar] 2. Баркер Дж, Хамфри Т.Дж., Браун М.В.Выживание Escherichia coli O157 в почвенных простейших: последствия для болезней. FEMS Microbiol Lett. 1999; 173: 291–295. [PubMed] [Google Scholar] 3. Барретт Т.Дж., Лиор Х., Грин Дж. Х., Хахрия Р., Уэллс Дж. Г., Белл Б. П., Грин К. Д., Льюис Дж., Гриффин П. М.. Лабораторное исследование мультистатной пищевой вспышки Escherichia coli O157: H7 с использованием гель-электрофореза в импульсном поле и фагового типирования. J Clin Microbiol. 1994; 32: 3013–3017. [Бесплатная статья PMC] [PubMed] [Google Scholar] 4.Бауэр М.Э., Велч Р.А. Характеристика токсина RTX от энтерогеморрагической Escherichia coli O157: H7. Заражение иммунной. 1996. 64: 167–175. [Бесплатная статья PMC] [PubMed] [Google Scholar] 6. Бурлин П., МакИвен С.А., Бурлин-Петцольд Ф., Уилсон Дж. Б., Джонсон Р.П., Джайлз К.Л. Связь между факторами вирулентности шига-токсина Escherichia coli и болезнью человека. J Clin Microbiol. 1999; 37: 497–503. [Бесплатная статья PMC] [PubMed] [Google Scholar] 7. Brashears MM, Galyean ML, Loneragan GH, Mann JE, Killinger-Mann K.Распространенность Escherichia coli O157: H7 и продуктивность мясного откорма крупного рогатого скота, получавшего Lactobacillus микробов прямого кормления. J Food Prot. 2003. 66: 748–754. [PubMed] [Google Scholar] 8. Брундер В., Шмидт Х., Карч Х. EspP, новая внеклеточная сериновая протеаза энтерогеморрагической Escherichia coli O157: H7 расщепляет человеческий фактор свертывания крови V.
J Clin Microbiol. 1994; 32: 3013–3017. [Бесплатная статья PMC] [PubMed] [Google Scholar] 4.Бауэр М.Э., Велч Р.А. Характеристика токсина RTX от энтерогеморрагической Escherichia coli O157: H7. Заражение иммунной. 1996. 64: 167–175. [Бесплатная статья PMC] [PubMed] [Google Scholar] 6. Бурлин П., МакИвен С.А., Бурлин-Петцольд Ф., Уилсон Дж. Б., Джонсон Р.П., Джайлз К.Л. Связь между факторами вирулентности шига-токсина Escherichia coli и болезнью человека. J Clin Microbiol. 1999; 37: 497–503. [Бесплатная статья PMC] [PubMed] [Google Scholar] 7. Brashears MM, Galyean ML, Loneragan GH, Mann JE, Killinger-Mann K.Распространенность Escherichia coli O157: H7 и продуктивность мясного откорма крупного рогатого скота, получавшего Lactobacillus микробов прямого кормления. J Food Prot. 2003. 66: 748–754. [PubMed] [Google Scholar] 8. Брундер В., Шмидт Х., Карч Х. EspP, новая внеклеточная сериновая протеаза энтерогеморрагической Escherichia coli O157: H7 расщепляет человеческий фактор свертывания крови V. Mol Microbiol. 1997; 24: 767–778. [PubMed] [Google Scholar] 9. Брундер В., Шмидт Х., Карч Х. KatP, новая каталаза-пероксидаза, кодируемая большой плазмидой энтерогеморрагической Escherichia coli 0157: H7.Микробиология. 1996. 142: 3305–3315. [PubMed] [Google Scholar] 10. Берланд В., Шао Ю., Перна Н. Т., Планкетт Г., София Х. Дж., Блаттнер Фр. Полная последовательность ДНК и анализ большой плазмиды вирулентности Escherichia coli O157: H7. Nucl Acids Res. 1998. 26: 4196–4204. [Бесплатная статья PMC] [PubMed] [Google Scholar] 11. Caprioli A, Morabito S, Brugère H, Oswald E. Enterohaemorrhagic Escherichia coli : Новые вопросы о вирулентности и способах передачи. Vet Res. 2005; 36: 289–311.[PubMed] [Google Scholar] 12. Кастани-Корнет член парламента, Пенфаунд Т.А., Смит Д., Эллиотт Дж. Ф., Фостер Дж. У. Контроль кислотостойкости Escherichia coli . J Bacteriol. 1999. 181: 3525–3535. [Бесплатная статья PMC] [PubMed] [Google Scholar] 13. Чо С., Бендер Дж.
Mol Microbiol. 1997; 24: 767–778. [PubMed] [Google Scholar] 9. Брундер В., Шмидт Х., Карч Х. KatP, новая каталаза-пероксидаза, кодируемая большой плазмидой энтерогеморрагической Escherichia coli 0157: H7.Микробиология. 1996. 142: 3305–3315. [PubMed] [Google Scholar] 10. Берланд В., Шао Ю., Перна Н. Т., Планкетт Г., София Х. Дж., Блаттнер Фр. Полная последовательность ДНК и анализ большой плазмиды вирулентности Escherichia coli O157: H7. Nucl Acids Res. 1998. 26: 4196–4204. [Бесплатная статья PMC] [PubMed] [Google Scholar] 11. Caprioli A, Morabito S, Brugère H, Oswald E. Enterohaemorrhagic Escherichia coli : Новые вопросы о вирулентности и способах передачи. Vet Res. 2005; 36: 289–311.[PubMed] [Google Scholar] 12. Кастани-Корнет член парламента, Пенфаунд Т.А., Смит Д., Эллиотт Дж. Ф., Фостер Дж. У. Контроль кислотостойкости Escherichia coli . J Bacteriol. 1999. 181: 3525–3535. [Бесплатная статья PMC] [PubMed] [Google Scholar] 13. Чо С., Бендер Дж. Б., Диез-Гонсалес Ф., Фосслер С. П., Хедберг С. В., Канини Дж. Б., Рюгг П. Л., Варник Л. Д., Уэллс С. Дж.. Распространенность и характеристика изолятов Escherichia coli O157 с молочных ферм Миннесоты и окружных ярмарок. J Food Prot. 2006; 69: 252–259. [PubMed] [Google Scholar] 14.Cray WC, Jr, Moon HW. Экспериментальное заражение телят и взрослого крупного рогатого скота Escherichia coli O157: H7. Appl Environ Microbiol. 1995; 61: 1586–1590. [Бесплатная статья PMC] [PubMed] [Google Scholar] 15. Delahay RM, Frankel G, Knutton S. Интимные взаимодействия энтеропатогенных Escherichia coli на поверхности клетки-хозяина. Curr Opin Infect Dis. 2001. 14: 559–565. [PubMed] [Google Scholar] 16. Дэн В., Пуэнте Дж. Л., Грюнхайд С., Ли Ю., Валланс Б. А., Васкес А. и др. Рассекающая вирулентность: систематический и функциональный анализ острова патогенности.Proc Natl Acad Sci USA. 2004. 101: 3597–3602. [Бесплатная статья PMC] [PubMed] [Google Scholar] 17. Добриндт Ю., Агерер Ф.
Б., Диез-Гонсалес Ф., Фосслер С. П., Хедберг С. В., Канини Дж. Б., Рюгг П. Л., Варник Л. Д., Уэллс С. Дж.. Распространенность и характеристика изолятов Escherichia coli O157 с молочных ферм Миннесоты и окружных ярмарок. J Food Prot. 2006; 69: 252–259. [PubMed] [Google Scholar] 14.Cray WC, Jr, Moon HW. Экспериментальное заражение телят и взрослого крупного рогатого скота Escherichia coli O157: H7. Appl Environ Microbiol. 1995; 61: 1586–1590. [Бесплатная статья PMC] [PubMed] [Google Scholar] 15. Delahay RM, Frankel G, Knutton S. Интимные взаимодействия энтеропатогенных Escherichia coli на поверхности клетки-хозяина. Curr Opin Infect Dis. 2001. 14: 559–565. [PubMed] [Google Scholar] 16. Дэн В., Пуэнте Дж. Л., Грюнхайд С., Ли Ю., Валланс Б. А., Васкес А. и др. Рассекающая вирулентность: систематический и функциональный анализ острова патогенности.Proc Natl Acad Sci USA. 2004. 101: 3597–3602. [Бесплатная статья PMC] [PubMed] [Google Scholar] 17. Добриндт Ю., Агерер Ф. , Михаэлис К., Янка А., Бухризер С., Самуэльсон М. и др. Анализ пластичности генома патогенных и комменсальных изолятов Escherichia coli с использованием массивов ДНК. J Bacteriol. 2003; 185: 1831–1840. [Бесплатная статья PMC] [PubMed] [Google Scholar] 18. Данн Дж. Р., Кин Дж. Э., Томпсон Р. А.. Распространенность Shiga-toxigenic Escherichia coli O157: H7 у взрослого молочного скота. J Am Vet Med Assoc.2004. 224: 1151–1158. [PubMed] [Google Scholar] 19. Дзива Ф., Махаджан А., Кэмерон П., Карри С., МакКендрик И.Дж., Уоллис Т.С., Смит Д.Г., Стивенс М.П. EspP, сериновая протеаза энтерогеморрагической Escherichia coli O157: H7, секретируемая типом V, влияет на колонизацию кишечника телят и прикрепление к первичным кишечным эпителиальным клеткам крупного рогатого скота. FEMS Microbiol Lett. 2007. 271: 258–264. [PubMed] [Google Scholar] 20. Эллиотт С.Дж., Сперандио В., Хирон Д.А., Шин С., Меллис Д.Л., Уэйнрайт Л., Хатчесон С.В., МакДэниел Т.К., Капер Дж.Б.Регулятор, кодируемый локусом сглаживания энтероцитов (LEE), контролирует экспрессию как LEE-, так и не-LEE-кодируемых факторов вирулентности в энтеропатогенных и энтерогеморрагических Escherichia coli .
, Михаэлис К., Янка А., Бухризер С., Самуэльсон М. и др. Анализ пластичности генома патогенных и комменсальных изолятов Escherichia coli с использованием массивов ДНК. J Bacteriol. 2003; 185: 1831–1840. [Бесплатная статья PMC] [PubMed] [Google Scholar] 18. Данн Дж. Р., Кин Дж. Э., Томпсон Р. А.. Распространенность Shiga-toxigenic Escherichia coli O157: H7 у взрослого молочного скота. J Am Vet Med Assoc.2004. 224: 1151–1158. [PubMed] [Google Scholar] 19. Дзива Ф., Махаджан А., Кэмерон П., Карри С., МакКендрик И.Дж., Уоллис Т.С., Смит Д.Г., Стивенс М.П. EspP, сериновая протеаза энтерогеморрагической Escherichia coli O157: H7, секретируемая типом V, влияет на колонизацию кишечника телят и прикрепление к первичным кишечным эпителиальным клеткам крупного рогатого скота. FEMS Microbiol Lett. 2007. 271: 258–264. [PubMed] [Google Scholar] 20. Эллиотт С.Дж., Сперандио В., Хирон Д.А., Шин С., Меллис Д.Л., Уэйнрайт Л., Хатчесон С.В., МакДэниел Т.К., Капер Дж.Б.Регулятор, кодируемый локусом сглаживания энтероцитов (LEE), контролирует экспрессию как LEE-, так и не-LEE-кодируемых факторов вирулентности в энтеропатогенных и энтерогеморрагических Escherichia coli . Заражение иммунной. 2000. 68: 6115–6126. [Бесплатная статья PMC] [PubMed] [Google Scholar] 21. Frenzen PD, Drake A, Angulo FJ. Экономическая стоимость болезни, вызванной инфекциями, вызванными Escherichia coli, O157 в США. J Food Prot. 2005; 68: 2623–2630. [PubMed] [Google Scholar] 22. Frost LS, Leplae R, Summers AO, Toussaint A.Мобильные генетические элементы: агенты эволюции открытого исходного кода. Nat Rev Microbiol. 2005; 3: 722–732. [PubMed] [Google Scholar] 23. Ганшерофф Л.Дж., О’Брайен А.Д. Escherichia coli O157: H7 у мясного скота, отправленного на убой в США: более высокие показатели распространенности, чем предполагалось ранее. Proc Natl Acad Sci USA. 2000; 97: 2959–2961. [Бесплатная статья PMC] [PubMed] [Google Scholar] 24. Гриффин PM, Tauxe RV. Эпидемиология инфекций, вызываемых Escherichia coli O157: H7, другими энтерогеморрагическими E.coli и связанный с ним гемолитико-уремический синдром. Epidemiol Rev.1991; 13: 60–98. [PubMed] [Google Scholar] 25.
Заражение иммунной. 2000. 68: 6115–6126. [Бесплатная статья PMC] [PubMed] [Google Scholar] 21. Frenzen PD, Drake A, Angulo FJ. Экономическая стоимость болезни, вызванной инфекциями, вызванными Escherichia coli, O157 в США. J Food Prot. 2005; 68: 2623–2630. [PubMed] [Google Scholar] 22. Frost LS, Leplae R, Summers AO, Toussaint A.Мобильные генетические элементы: агенты эволюции открытого исходного кода. Nat Rev Microbiol. 2005; 3: 722–732. [PubMed] [Google Scholar] 23. Ганшерофф Л.Дж., О’Брайен А.Д. Escherichia coli O157: H7 у мясного скота, отправленного на убой в США: более высокие показатели распространенности, чем предполагалось ранее. Proc Natl Acad Sci USA. 2000; 97: 2959–2961. [Бесплатная статья PMC] [PubMed] [Google Scholar] 24. Гриффин PM, Tauxe RV. Эпидемиология инфекций, вызываемых Escherichia coli O157: H7, другими энтерогеморрагическими E.coli и связанный с ним гемолитико-уремический синдром. Epidemiol Rev.1991; 13: 60–98. [PubMed] [Google Scholar] 25. Грис Т.Э., Сигель М.Б., Lathem WW, Велч Р.А. Протеаза StcE способствует тесному прилипанию энтерогеморрагической Escherichia coli O157: H7 к клеткам-хозяевам. Заражение иммунной. 2005. 73: 1295–1303. [Бесплатная статья PMC] [PubMed] [Google Scholar] 26. Хейлз Б.А., Харт Калифорния, Батт Р.М., Сондерс-младший. Большие плазмиды, обнаруженные в энтерогеморрагической и энтеропатогенной Escherichia coli , составляют родственную серию репликонов Inc F-IIA, дефектных по переносу.Плазмида. 1992; 28: 183–193. [PubMed] [Google Scholar] 27. Хэнкок Д.Д., Бессер Т.Э., Райс Д.Х., Херриотт Д.Е., Тарр П.И. Продольное исследование Escherichia coli O157 в четырнадцати стадах крупного рогатого скота. Epidemiol Infect. 1997. 118: 193–195. [Бесплатная статья PMC] [PubMed] [Google Scholar] 28. Heuvelink AE, van Heerwaarden C, Zwartkruis-Nahuis JT, van Oosterom R, Edink K, van Duynhoven YT, de Boer E. Escherichia coli O157 Инфекция, связанная с контактным зоопарком.
Грис Т.Э., Сигель М.Б., Lathem WW, Велч Р.А. Протеаза StcE способствует тесному прилипанию энтерогеморрагической Escherichia coli O157: H7 к клеткам-хозяевам. Заражение иммунной. 2005. 73: 1295–1303. [Бесплатная статья PMC] [PubMed] [Google Scholar] 26. Хейлз Б.А., Харт Калифорния, Батт Р.М., Сондерс-младший. Большие плазмиды, обнаруженные в энтерогеморрагической и энтеропатогенной Escherichia coli , составляют родственную серию репликонов Inc F-IIA, дефектных по переносу.Плазмида. 1992; 28: 183–193. [PubMed] [Google Scholar] 27. Хэнкок Д.Д., Бессер Т.Э., Райс Д.Х., Херриотт Д.Е., Тарр П.И. Продольное исследование Escherichia coli O157 в четырнадцати стадах крупного рогатого скота. Epidemiol Infect. 1997. 118: 193–195. [Бесплатная статья PMC] [PubMed] [Google Scholar] 28. Heuvelink AE, van Heerwaarden C, Zwartkruis-Nahuis JT, van Oosterom R, Edink K, van Duynhoven YT, de Boer E. Escherichia coli O157 Инфекция, связанная с контактным зоопарком. Epidemiol Infect. 2002; 129: 295–302.[Бесплатная статья PMC] [PubMed] [Google Scholar] 29. Яцевич М.С., Ачесон Д.В., Binion DG, West GA, Lincicome LL, Fiocchi C, Keusch GT. Ответы эндотелиальных клеток микрососудов кишечника человека на токсины шига 1 и 2 и патогенез геморрагического колита. Заражение иммунной. 1999; 67: 1439–1444. [Бесплатная статья PMC] [PubMed] [Google Scholar] 31. Джонсон Р.П., Уилсон Дж. Б., Мишель П., Ран К., Ренвик С. А., Джайлз К. Л., Спика Дж. С.. Заражение человека веротоксигенной Escherichia coli связано с воздействием на фермы и сельскую среду.В: Стюарт К.С., Флинтс Х.Дж., редакторы. Escherichia coli O157 в сельскохозяйственных животных. CABI Publishing; Уоллингфорд, Великобритания: 1999. С. 147–168. [Google Scholar] 32. Канюк Н.А., Виноградов Э., Ли Дж., Монтейро М.А., Уитфилд С. Ферменты, кодируемые хромосомами и плазмидами, необходимы для сборки центрального олигосахарида типа R 3 в липополисахариде Escherichia coli O157: H7. J Biol Chem.
Epidemiol Infect. 2002; 129: 295–302.[Бесплатная статья PMC] [PubMed] [Google Scholar] 29. Яцевич М.С., Ачесон Д.В., Binion DG, West GA, Lincicome LL, Fiocchi C, Keusch GT. Ответы эндотелиальных клеток микрососудов кишечника человека на токсины шига 1 и 2 и патогенез геморрагического колита. Заражение иммунной. 1999; 67: 1439–1444. [Бесплатная статья PMC] [PubMed] [Google Scholar] 31. Джонсон Р.П., Уилсон Дж. Б., Мишель П., Ран К., Ренвик С. А., Джайлз К. Л., Спика Дж. С.. Заражение человека веротоксигенной Escherichia coli связано с воздействием на фермы и сельскую среду.В: Стюарт К.С., Флинтс Х.Дж., редакторы. Escherichia coli O157 в сельскохозяйственных животных. CABI Publishing; Уоллингфорд, Великобритания: 1999. С. 147–168. [Google Scholar] 32. Канюк Н.А., Виноградов Э., Ли Дж., Монтейро М.А., Уитфилд С. Ферменты, кодируемые хромосомами и плазмидами, необходимы для сборки центрального олигосахарида типа R 3 в липополисахариде Escherichia coli O157: H7. J Biol Chem.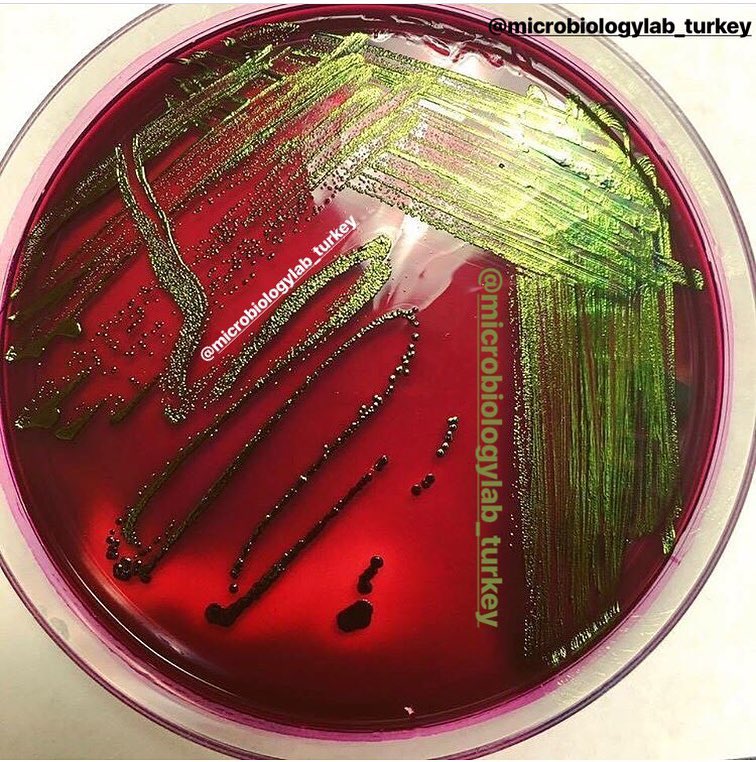 2004. 279: 31237–31250. [PubMed] [Google Scholar] 33. Капер Дж. Б., Натаро Дж. П., Мобли Х.Л. Патогенный Escherichia coli .Nat Rev Microbiol. 2004; 2: 123–140. [PubMed] [Google Scholar] 34. Кармали М.А., Стил Б.Т., Петрик М., Лим С. Спорадические случаи гемолитико-уремического синдрома, связанные с фекальным цитотоксином и продуцирующим цитотоксин Escherichia coli в стуле. Ланцет. 1983; 1: 619–620. [PubMed] [Google Scholar] 35. Kim SH, Jia W, Bishop RE, Gyles C. Гомолог msbB, содержащийся в плазмиде pO157, кодирует ацилтрансферазу, участвующую в биосинтезе липида A в Escherichia coli O157: H7. Заражение иммунной. 2004. 72: 1174–1180.[Бесплатная статья PMC] [PubMed] [Google Scholar] 36. Klapproth JMA, I, Scaletsky CA, McNamara BP, Lai LC, Malstrom C, James SP, Donnenberg MS. Большой токсин патогенного штамма Escherichia coli , подавляющий активацию лимфоцитов. Заражение иммунной. 2000. 68: 2148–2155. [Бесплатная статья PMC] [PubMed] [Google Scholar] 37.
2004. 279: 31237–31250. [PubMed] [Google Scholar] 33. Капер Дж. Б., Натаро Дж. П., Мобли Х.Л. Патогенный Escherichia coli .Nat Rev Microbiol. 2004; 2: 123–140. [PubMed] [Google Scholar] 34. Кармали М.А., Стил Б.Т., Петрик М., Лим С. Спорадические случаи гемолитико-уремического синдрома, связанные с фекальным цитотоксином и продуцирующим цитотоксин Escherichia coli в стуле. Ланцет. 1983; 1: 619–620. [PubMed] [Google Scholar] 35. Kim SH, Jia W, Bishop RE, Gyles C. Гомолог msbB, содержащийся в плазмиде pO157, кодирует ацилтрансферазу, участвующую в биосинтезе липида A в Escherichia coli O157: H7. Заражение иммунной. 2004. 72: 1174–1180.[Бесплатная статья PMC] [PubMed] [Google Scholar] 36. Klapproth JMA, I, Scaletsky CA, McNamara BP, Lai LC, Malstrom C, James SP, Donnenberg MS. Большой токсин патогенного штамма Escherichia coli , подавляющий активацию лимфоцитов. Заражение иммунной. 2000. 68: 2148–2155. [Бесплатная статья PMC] [PubMed] [Google Scholar] 37.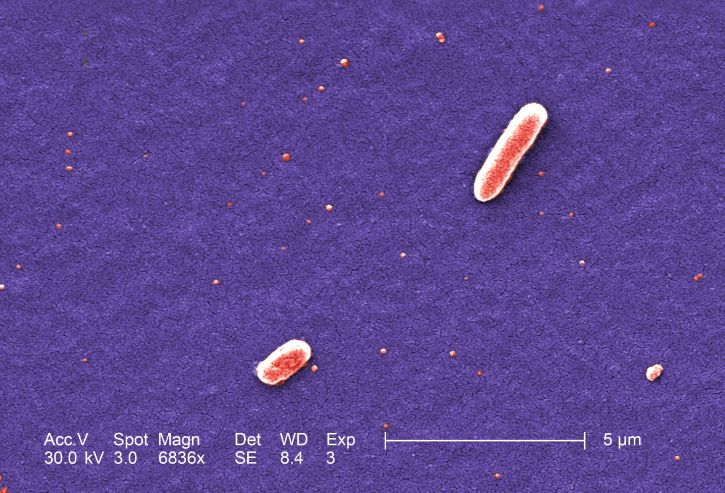 Lathem WW, Grys TE, Witowski SE, Torres AG, Kaper JB, Tarr PI, Welch RA. StcE, металлопротеаза, секретируемая Escherichia coli O157: H7, специфически расщепляет ингибитор C1-эстеразы.Mol Microbiol. 2002. 45: 277–288. [PubMed] [Google Scholar] 38. LeJeune JT, Besser TE, Hancock DD. Поилки для крупного рогатого скота как резервуары для Escherichia coli O157. Appl Environ Microbiol. 2001; 67: 3053–3057. [Бесплатная статья PMC] [PubMed] [Google Scholar] 39. Лим Дж.Й., Ли Дж., Шенг Х., Бессер Т.Э., Поттер К., Ховде С.Дж. Escherichia coli O157: Колонизация H7 в прямой кишке крупного рогатого скота с длительным культивированием. Appl Environ Microbiol. 2007. 73: 1380–1382. [Бесплатная статья PMC] [PubMed] [Google Scholar] 40.Лим Дж.Й., Шенг Х., Со К.С., Пак Й.Х., Ховде СиДжей. Характеристика мутанта с делецией O157 плазмиды Escherichia coli O157: H7, а также его выживаемость и устойчивость у крупного рогатого скота. Appl Environ Microbiol. 2007. 73: 2037–2047.
Lathem WW, Grys TE, Witowski SE, Torres AG, Kaper JB, Tarr PI, Welch RA. StcE, металлопротеаза, секретируемая Escherichia coli O157: H7, специфически расщепляет ингибитор C1-эстеразы.Mol Microbiol. 2002. 45: 277–288. [PubMed] [Google Scholar] 38. LeJeune JT, Besser TE, Hancock DD. Поилки для крупного рогатого скота как резервуары для Escherichia coli O157. Appl Environ Microbiol. 2001; 67: 3053–3057. [Бесплатная статья PMC] [PubMed] [Google Scholar] 39. Лим Дж.Й., Ли Дж., Шенг Х., Бессер Т.Э., Поттер К., Ховде С.Дж. Escherichia coli O157: Колонизация H7 в прямой кишке крупного рогатого скота с длительным культивированием. Appl Environ Microbiol. 2007. 73: 1380–1382. [Бесплатная статья PMC] [PubMed] [Google Scholar] 40.Лим Дж.Й., Шенг Х., Со К.С., Пак Й.Х., Ховде СиДжей. Характеристика мутанта с делецией O157 плазмиды Escherichia coli O157: H7, а также его выживаемость и устойчивость у крупного рогатого скота. Appl Environ Microbiol. 2007. 73: 2037–2047. [Бесплатная статья PMC] [PubMed] [Google Scholar] 41. Макино К., Исии К., Ясунага Т., Хаттори М., Йокояма К., Юцудо С.Х. и др. Полные нуклеотидные последовательности плазмид размером 93 т.п.н. и 3,3 т.п.н. энтерогеморрагической Escherichia coli O157: H7, полученной из очага Сакаи. ДНК Res.1998; 5: 1–9. [PubMed] [Google Scholar] 42. Мауле А. Выживание вероцитотоксигенных Escherichia coli O157 в почве, воде и на поверхностях. Symp Ser Soc Appl Microbiol. 2000; 29: 71С – 78С. [PubMed] [Google Scholar] 43. Мид П.С., Слуцкер Л., Дитц В., Маккейг Л.Ф., Брези Дж.С., Шапиро С., Гриффин П.М., Токс Р.В. Заболевания и смерть, связанные с пищевыми продуктами в США. Emerg Infect Dis. 1999; 5: 607–625. [Бесплатная статья PMC] [PubMed] [Google Scholar] 44. Мелтон-Селса АР, Дарнелл СК, О’Брайен А.Д. Активация Shiga-подобных токсинов кишечной слизью мышей и человека коррелирует с вирулентностью энтерогеморрагических изолятов Escherichia coli O91: h31 у перорально инфицированных мышей, получавших стрептомицин.
[Бесплатная статья PMC] [PubMed] [Google Scholar] 41. Макино К., Исии К., Ясунага Т., Хаттори М., Йокояма К., Юцудо С.Х. и др. Полные нуклеотидные последовательности плазмид размером 93 т.п.н. и 3,3 т.п.н. энтерогеморрагической Escherichia coli O157: H7, полученной из очага Сакаи. ДНК Res.1998; 5: 1–9. [PubMed] [Google Scholar] 42. Мауле А. Выживание вероцитотоксигенных Escherichia coli O157 в почве, воде и на поверхностях. Symp Ser Soc Appl Microbiol. 2000; 29: 71С – 78С. [PubMed] [Google Scholar] 43. Мид П.С., Слуцкер Л., Дитц В., Маккейг Л.Ф., Брези Дж.С., Шапиро С., Гриффин П.М., Токс Р.В. Заболевания и смерть, связанные с пищевыми продуктами в США. Emerg Infect Dis. 1999; 5: 607–625. [Бесплатная статья PMC] [PubMed] [Google Scholar] 44. Мелтон-Селса АР, Дарнелл СК, О’Брайен А.Д. Активация Shiga-подобных токсинов кишечной слизью мышей и человека коррелирует с вирулентностью энтерогеморрагических изолятов Escherichia coli O91: h31 у перорально инфицированных мышей, получавших стрептомицин. Заражение иммунной. 1996. 64: 1569–1576. [Бесплатная статья PMC] [PubMed] [Google Scholar] 45. Мичино Х, Араки К., Минами С., Такая С., Сакаи Н., Миядзаки М., Оно А., Янагава Х. Массовая вспышка Escherichia coli O157: инфекция H7 у школьников в городе Сакаи, Япония, связанная с потреблением белого редиса ростки. Am J Epidemiol. 1999; 150: 787–796. [PubMed] [Google Scholar] 46. Морабито С., Тоццоли Р., Освальд Е., Каприоли А. Мозаичный остров патогенности, состоящий из локуса сглаживания энтероцитов и острова патогенности Escherichia coli O157: H7 часто присутствует в прикреплении и удалении E.coli . Заражение иммунной. 2003. 71: 3343–3348. [Бесплатная статья PMC] [PubMed] [Google Scholar] 48. Naylor SW, Low JC, Besser TE, Mahajan A, Gunn GJ, Pearce MC, McKendrick IJ, Smith DGE, Gally DL. Плотная слизистая оболочка лимфоидного фолликула в терминальной части прямой кишки является основным местом колонизации энтерогеморрагической Escherichia coli O157: H7 в организме крупного рогатого скота.
Заражение иммунной. 1996. 64: 1569–1576. [Бесплатная статья PMC] [PubMed] [Google Scholar] 45. Мичино Х, Араки К., Минами С., Такая С., Сакаи Н., Миядзаки М., Оно А., Янагава Х. Массовая вспышка Escherichia coli O157: инфекция H7 у школьников в городе Сакаи, Япония, связанная с потреблением белого редиса ростки. Am J Epidemiol. 1999; 150: 787–796. [PubMed] [Google Scholar] 46. Морабито С., Тоццоли Р., Освальд Е., Каприоли А. Мозаичный остров патогенности, состоящий из локуса сглаживания энтероцитов и острова патогенности Escherichia coli O157: H7 часто присутствует в прикреплении и удалении E.coli . Заражение иммунной. 2003. 71: 3343–3348. [Бесплатная статья PMC] [PubMed] [Google Scholar] 48. Naylor SW, Low JC, Besser TE, Mahajan A, Gunn GJ, Pearce MC, McKendrick IJ, Smith DGE, Gally DL. Плотная слизистая оболочка лимфоидного фолликула в терминальной части прямой кишки является основным местом колонизации энтерогеморрагической Escherichia coli O157: H7 в организме крупного рогатого скота.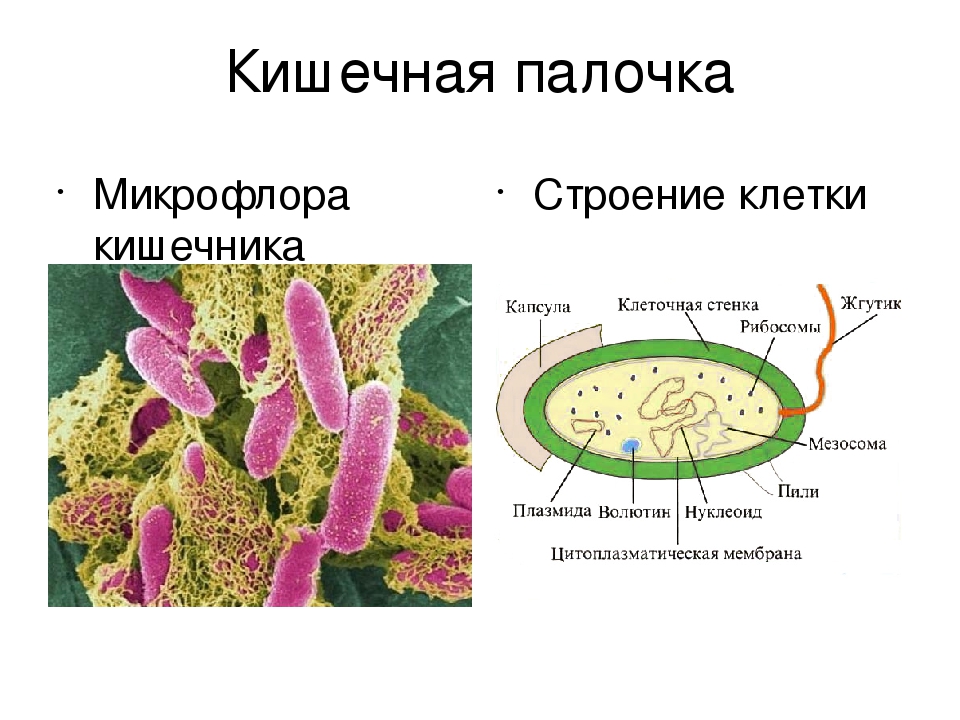 Заражение иммунной. 2003. 71: 1505–1512. [Бесплатная статья PMC] [PubMed] [Google Scholar] 49. Orndorff PE, Wang Y, Huang SH, Wass CA, Stins MF, Kim KS.Локус гена yijP способствует инвазии Escherichia coli K1 в эндотелиальные клетки микрососудов головного мозга. Заражение иммунной. 1999; 67: 4751–4756. [Бесплатная статья PMC] [PubMed] [Google Scholar] 50. Острофф С.М., Тарр П.И., Нил М.А., Льюис Дж. Х., Харгретт-Бин Н., Кобаяши Дж. М.. Генотипы токсинов и плазмидные профили как детерминанты системных осложнений при инфекциях Escherichia coli O157: H7. J Infect Dis. 1989; 160: 994–998. [PubMed] [Google Scholar] 51. Патон А.В., Патон Дж.С. Прямое обнаружение Shiga токсигенных штаммов Escherichia coli , принадлежащих к серогруппам O111, O157 и O113, методом мультиплексной ПЦР.J Clin Microbiol. 1999; 37: 3362–3365. [Бесплатная статья PMC] [PubMed] [Google Scholar] 52. Perna NT, Mayhew GF, Posfai G, Elliott S, Donnenberg MS, Kaper JB, Blattner FR. Молекулярная эволюция острова патогенности от энтерогеморрагической Escherichia coli O157: H7.
Заражение иммунной. 2003. 71: 1505–1512. [Бесплатная статья PMC] [PubMed] [Google Scholar] 49. Orndorff PE, Wang Y, Huang SH, Wass CA, Stins MF, Kim KS.Локус гена yijP способствует инвазии Escherichia coli K1 в эндотелиальные клетки микрососудов головного мозга. Заражение иммунной. 1999; 67: 4751–4756. [Бесплатная статья PMC] [PubMed] [Google Scholar] 50. Острофф С.М., Тарр П.И., Нил М.А., Льюис Дж. Х., Харгретт-Бин Н., Кобаяши Дж. М.. Генотипы токсинов и плазмидные профили как детерминанты системных осложнений при инфекциях Escherichia coli O157: H7. J Infect Dis. 1989; 160: 994–998. [PubMed] [Google Scholar] 51. Патон А.В., Патон Дж.С. Прямое обнаружение Shiga токсигенных штаммов Escherichia coli , принадлежащих к серогруппам O111, O157 и O113, методом мультиплексной ПЦР.J Clin Microbiol. 1999; 37: 3362–3365. [Бесплатная статья PMC] [PubMed] [Google Scholar] 52. Perna NT, Mayhew GF, Posfai G, Elliott S, Donnenberg MS, Kaper JB, Blattner FR. Молекулярная эволюция острова патогенности от энтерогеморрагической Escherichia coli O157: H7. Заражение иммунной. 1998; 66: 3810–3817. [Бесплатная статья PMC] [PubMed] [Google Scholar] 53. Perna NT, Plunkett G, Burland V, Mau B, Glasner JD, Rose DJ и др. Последовательность генома энтерогеморрагической Escherichia coli O157: H7.Природа. 2001; 409: 529–533. [PubMed] [Google Scholar] 54. Петерсон В.Л., Макковяк П.А., Барнетт С.С., Марлинг-Кейсон М., Хейли М.Л. Бактерицидный барьер желудка человека: механизмы действия, относительная антибактериальная активность и влияние питания. J Infect Dis. 1989; 159: 979–983. [PubMed] [Google Scholar] 55. Putonti C, Luo Y, Katili C, Chumakov S, Fox GE, Graur D, Fofanov Y. Вычислительный инструмент для геномной идентификации регионов с необычными композиционными свойствами и его использование при обнаружении горизонтально перенесенных последовательностей.Mol Biol Evol. 2006; 23: 1863–1868. [PubMed] [Google Scholar] 56. Ранжел Дж. М., Спарлинг П. Х., Кроу К., Гриффин П. М., Свердлов Д. Л.. Эпидемиология вспышек Escherichia coli O157: H7, США, 1982–2002 гг.
Заражение иммунной. 1998; 66: 3810–3817. [Бесплатная статья PMC] [PubMed] [Google Scholar] 53. Perna NT, Plunkett G, Burland V, Mau B, Glasner JD, Rose DJ и др. Последовательность генома энтерогеморрагической Escherichia coli O157: H7.Природа. 2001; 409: 529–533. [PubMed] [Google Scholar] 54. Петерсон В.Л., Макковяк П.А., Барнетт С.С., Марлинг-Кейсон М., Хейли М.Л. Бактерицидный барьер желудка человека: механизмы действия, относительная антибактериальная активность и влияние питания. J Infect Dis. 1989; 159: 979–983. [PubMed] [Google Scholar] 55. Putonti C, Luo Y, Katili C, Chumakov S, Fox GE, Graur D, Fofanov Y. Вычислительный инструмент для геномной идентификации регионов с необычными композиционными свойствами и его использование при обнаружении горизонтально перенесенных последовательностей.Mol Biol Evol. 2006; 23: 1863–1868. [PubMed] [Google Scholar] 56. Ранжел Дж. М., Спарлинг П. Х., Кроу К., Гриффин П. М., Свердлов Д. Л.. Эпидемиология вспышек Escherichia coli O157: H7, США, 1982–2002 гг.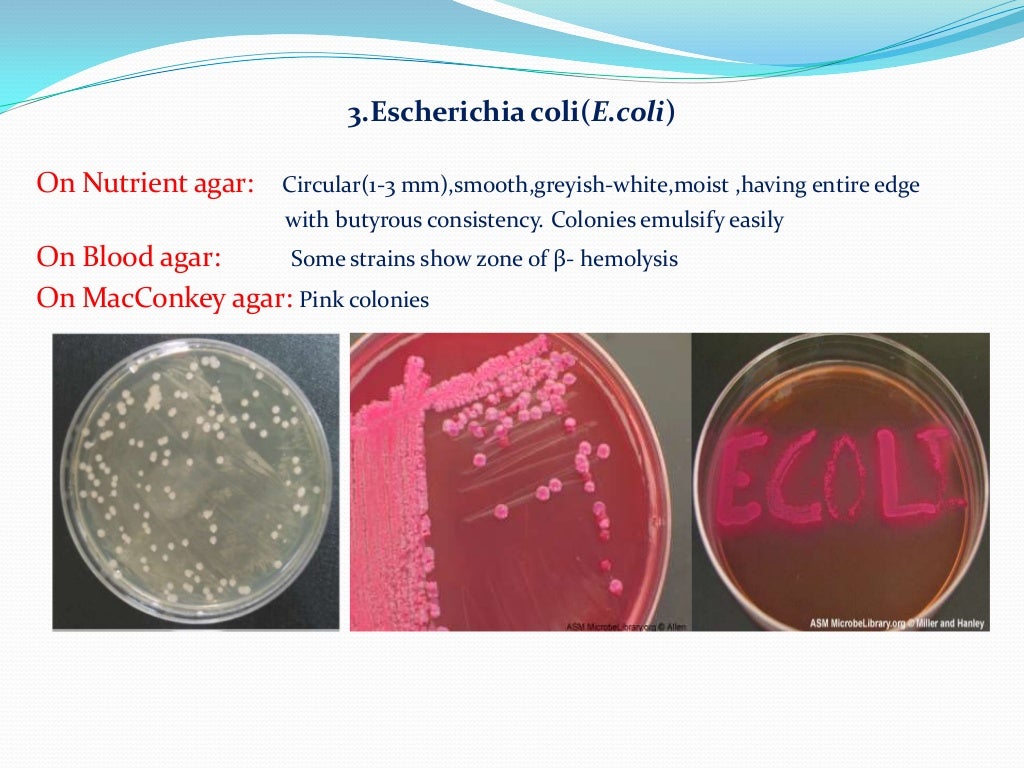 Emerg Infect Dis. 2005; 11: 603–609. [Бесплатная статья PMC] [PubMed] [Google Scholar] 57. Райли Л.У., Ремис Р.С., Хелгерсон С.Д., Макги Х.Б., Уэллс Дж. Г., Дэвис Б. Р. и др. Геморрагический колит, связанный с редким серотипом Escherichia coli . N Engl J Med. 1983; 308: 681–685. [PubMed] [Google Scholar] 58.Сандерсон М.В., Бессер Т.Э., Гей Дж.М., Гей CC, Хэнкок Д.Д. Фекал Escherichia coli O157: Характер выделения H7 у орально инокулированных телят. Vet Microbiol. 1999; 69: 199–205. [PubMed] [Google Scholar] 59. Саксена С.К., О’Брайен А.Д., Акерман Э.Дж. Токсин шига, вариант шига-подобного токсина II и рицин представляют собой односайтовую РНК N -гликозидазы 28S РНК при микроинъекции в ооциты Xenopus . J Biol Chem. 1989; 264: 596–601. [PubMed] [Google Scholar] 60. Шлех В.Ф., III, Чейз Д.П., Бэдли А.Модель пищевой инфекции Listeria monocytogenes у крыс Sprague-Dawley с использованием инокуляции желудка: развитие и влияние кислотности желудочного сока на инфекционную дозу.
Emerg Infect Dis. 2005; 11: 603–609. [Бесплатная статья PMC] [PubMed] [Google Scholar] 57. Райли Л.У., Ремис Р.С., Хелгерсон С.Д., Макги Х.Б., Уэллс Дж. Г., Дэвис Б. Р. и др. Геморрагический колит, связанный с редким серотипом Escherichia coli . N Engl J Med. 1983; 308: 681–685. [PubMed] [Google Scholar] 58.Сандерсон М.В., Бессер Т.Э., Гей Дж.М., Гей CC, Хэнкок Д.Д. Фекал Escherichia coli O157: Характер выделения H7 у орально инокулированных телят. Vet Microbiol. 1999; 69: 199–205. [PubMed] [Google Scholar] 59. Саксена С.К., О’Брайен А.Д., Акерман Э.Дж. Токсин шига, вариант шига-подобного токсина II и рицин представляют собой односайтовую РНК N -гликозидазы 28S РНК при микроинъекции в ооциты Xenopus . J Biol Chem. 1989; 264: 596–601. [PubMed] [Google Scholar] 60. Шлех В.Ф., III, Чейз Д.П., Бэдли А.Модель пищевой инфекции Listeria monocytogenes у крыс Sprague-Dawley с использованием инокуляции желудка: развитие и влияние кислотности желудочного сока на инфекционную дозу. Int J Food Microbiol. 1993; 18: 15–24. [PubMed] [Google Scholar] 61. Schmidt H, Beutin L, Karch H. Молекулярный анализ кодируемого плазмидой гемолизина Escherichia coli O157: штамм H7 EDL 933. Infect Immun. 1995; 63: 1055–1061. [Бесплатная статья PMC] [PubMed] [Google Scholar] 62. Schmidt H, Henkel B., Karch H. Кластер генов, тесно связанный с оперонами пути секреции типа II грамотрицательных бактерий, расположен на большой плазмиде энтерогеморрагических штаммов Escherichia coli O157.FEMS Microbiol Lett. 1997. 148: 265–272. [PubMed] [Google Scholar] 63. Шмидт Х., Карч Х., Бойтин Л. Большие плазмиды энтерогеморрагических штаммов Escherichia coli O157 кодируют гемолизины, которые предположительно являются членами семейства альфа-гемолизинов E. coli . FEMS Microbiol Lett. 1994; 117: 189–196. [PubMed] [Google Scholar] 64. Шенг Х., Лим Дж.Й., Кнехт Х.Д., Ли Дж., Ховде С.Дж. Роль факторов вирулентности Escherichia coli O157: H7 в колонизации терминальной слизистой оболочки прямой кишки крупного рогатого скота.
Int J Food Microbiol. 1993; 18: 15–24. [PubMed] [Google Scholar] 61. Schmidt H, Beutin L, Karch H. Молекулярный анализ кодируемого плазмидой гемолизина Escherichia coli O157: штамм H7 EDL 933. Infect Immun. 1995; 63: 1055–1061. [Бесплатная статья PMC] [PubMed] [Google Scholar] 62. Schmidt H, Henkel B., Karch H. Кластер генов, тесно связанный с оперонами пути секреции типа II грамотрицательных бактерий, расположен на большой плазмиде энтерогеморрагических штаммов Escherichia coli O157.FEMS Microbiol Lett. 1997. 148: 265–272. [PubMed] [Google Scholar] 63. Шмидт Х., Карч Х., Бойтин Л. Большие плазмиды энтерогеморрагических штаммов Escherichia coli O157 кодируют гемолизины, которые предположительно являются членами семейства альфа-гемолизинов E. coli . FEMS Microbiol Lett. 1994; 117: 189–196. [PubMed] [Google Scholar] 64. Шенг Х., Лим Дж.Й., Кнехт Х.Д., Ли Дж., Ховде С.Дж. Роль факторов вирулентности Escherichia coli O157: H7 в колонизации терминальной слизистой оболочки прямой кишки крупного рогатого скота. Заражение иммунной. 2006. 74: 4685–4693. [Бесплатная статья PMC] [PubMed] [Google Scholar] 65. Шима К., Йошии Н., Акиба М., Нисимура К., Наказава М., Ямасаки С. Сравнение ПЦР – ПДРФ и PFGE для определения клональности энтерогеморрагических штаммов Escherichia coli . FEMS Microbiol Lett. 2006; 257: 124–131. [PubMed] [Google Scholar] 66. Стивенс МП, Роу А.Дж., Влисиду И., Ван Димен П.М., Ла Раджионе Р.М., Лучший А, Вудворд М.Дж., Галли Д.Л., Уоллис Т.С. Мутация toxB и усеченная версия гена efa -1 в Escherichia coli O157: H7 влияет на экспрессию и секрецию локуса белков, кодируемых удалением энтероцитов, но не на колонизацию кишечника у телят или овец.Заражение иммунной. 2004. 72: 5402–5411. [Бесплатная статья PMC] [PubMed] [Google Scholar] 67. Tatsuno I, Horie M, Abe H, Miki T, Makino K, Shinagawa H, Taguchi H, Kamiya S, Hayashi T. ген toxB на pO157 энтерогеморрагической Escherichia coli O157: H7 необходим для полного фенотипа адгезии эпителиальных клеток .
Заражение иммунной. 2006. 74: 4685–4693. [Бесплатная статья PMC] [PubMed] [Google Scholar] 65. Шима К., Йошии Н., Акиба М., Нисимура К., Наказава М., Ямасаки С. Сравнение ПЦР – ПДРФ и PFGE для определения клональности энтерогеморрагических штаммов Escherichia coli . FEMS Microbiol Lett. 2006; 257: 124–131. [PubMed] [Google Scholar] 66. Стивенс МП, Роу А.Дж., Влисиду И., Ван Димен П.М., Ла Раджионе Р.М., Лучший А, Вудворд М.Дж., Галли Д.Л., Уоллис Т.С. Мутация toxB и усеченная версия гена efa -1 в Escherichia coli O157: H7 влияет на экспрессию и секрецию локуса белков, кодируемых удалением энтероцитов, но не на колонизацию кишечника у телят или овец.Заражение иммунной. 2004. 72: 5402–5411. [Бесплатная статья PMC] [PubMed] [Google Scholar] 67. Tatsuno I, Horie M, Abe H, Miki T, Makino K, Shinagawa H, Taguchi H, Kamiya S, Hayashi T. ген toxB на pO157 энтерогеморрагической Escherichia coli O157: H7 необходим для полного фенотипа адгезии эпителиальных клеток . Заражение иммунной. 2001; 69: 6660–6669. [Бесплатная статья PMC] [PubMed] [Google Scholar] 68. Томпсон Дж. С., Ходж Д. С., Борчик А. А.. Экспресс-биохимический тест для выявления вероцитотоксин-положительных штаммов Escherichia coli серотипа O157.J Clin Microbiol. 1990; 28: 2165–2168. [Бесплатная статья PMC] [PubMed] [Google Scholar] 69. ван Димен П.М., Дзива Ф., Стивенс МП, Уоллис Т.С. Идентификация энтерогеморрагической Escherichia coli O26: H-гены, необходимые для колонизации кишечника у телят. Заражение иммунной. 2005; 73: 1735–1743. [Бесплатная статья PMC] [PubMed] [Google Scholar] 70. Варма Дж. К., Грин К. Д., Реллер М. Э., Делонг С. М., Троттье Дж., Новицки С.Ф. и др. Вспышка инфекции Escherichia coli O157 в результате контакта с зараженным зданием.JAMA. 2003. 290: 2709–2712. [PubMed] [Google Scholar] 71. Уэллс Дж. Г., Дэвис Б. Р., Ваксмут И. К., Райли Л. В., Ремис Р. С., Соколов Р., Моррис Г. К.. Лабораторное исследование вспышек геморрагического колита, связанного с редким серотипом Escherichia coli .
Заражение иммунной. 2001; 69: 6660–6669. [Бесплатная статья PMC] [PubMed] [Google Scholar] 68. Томпсон Дж. С., Ходж Д. С., Борчик А. А.. Экспресс-биохимический тест для выявления вероцитотоксин-положительных штаммов Escherichia coli серотипа O157.J Clin Microbiol. 1990; 28: 2165–2168. [Бесплатная статья PMC] [PubMed] [Google Scholar] 69. ван Димен П.М., Дзива Ф., Стивенс МП, Уоллис Т.С. Идентификация энтерогеморрагической Escherichia coli O26: H-гены, необходимые для колонизации кишечника у телят. Заражение иммунной. 2005; 73: 1735–1743. [Бесплатная статья PMC] [PubMed] [Google Scholar] 70. Варма Дж. К., Грин К. Д., Реллер М. Э., Делонг С. М., Троттье Дж., Новицки С.Ф. и др. Вспышка инфекции Escherichia coli O157 в результате контакта с зараженным зданием.JAMA. 2003. 290: 2709–2712. [PubMed] [Google Scholar] 71. Уэллс Дж. Г., Дэвис Б. Р., Ваксмут И. К., Райли Л. В., Ремис Р. С., Соколов Р., Моррис Г. К.. Лабораторное исследование вспышек геморрагического колита, связанного с редким серотипом Escherichia coli . J Clin Microbiol. 1983; 18: 512–520. [Бесплатная статья PMC] [PubMed] [Google Scholar] 72. Вик Л.М., Ци В., Лачер Д.В., Уиттам Т.С. Эволюция геномного содержимого при ступенчатом появлении Escherichia coli O157: H7. J Bacteriol. 2005; 187: 1783–1791.[Бесплатная статья PMC] [PubMed] [Google Scholar] 73. Уиллшоу Г.А., Смит Х.Р., Чести Т., Уолл П.Г., Роу Б. Цитотоксин-продуцирующая вспышка Escherichia coli O157 в Англии и Уэльсе, 1995: Фенотипические методы и генотипическое определение подтипов. Emerg Infect Dis. 1997; 3: 561–565. [Бесплатная статья PMC] [PubMed] [Google Scholar] 75. Юн Дж.В., Лим Дж.Й., Пак Й.Х., Ховде СиДжей. Участие оперона Escherichia coli O157: H7 (pO157) ecf и активности миристоилтрансферазы липида А в выживаемости бактерий в желудочно-кишечном тракте крупного рогатого скота и устойчивости бактерий в поилках фермы.Заражение иммунной. 2005. 73: 2367–2378. [Бесплатная статья PMC] [PubMed] [Google Scholar] 76. Юн Дж. В., Миннич С.
J Clin Microbiol. 1983; 18: 512–520. [Бесплатная статья PMC] [PubMed] [Google Scholar] 72. Вик Л.М., Ци В., Лачер Д.В., Уиттам Т.С. Эволюция геномного содержимого при ступенчатом появлении Escherichia coli O157: H7. J Bacteriol. 2005; 187: 1783–1791.[Бесплатная статья PMC] [PubMed] [Google Scholar] 73. Уиллшоу Г.А., Смит Х.Р., Чести Т., Уолл П.Г., Роу Б. Цитотоксин-продуцирующая вспышка Escherichia coli O157 в Англии и Уэльсе, 1995: Фенотипические методы и генотипическое определение подтипов. Emerg Infect Dis. 1997; 3: 561–565. [Бесплатная статья PMC] [PubMed] [Google Scholar] 75. Юн Дж.В., Лим Дж.Й., Пак Й.Х., Ховде СиДжей. Участие оперона Escherichia coli O157: H7 (pO157) ecf и активности миристоилтрансферазы липида А в выживаемости бактерий в желудочно-кишечном тракте крупного рогатого скота и устойчивости бактерий в поилках фермы.Заражение иммунной. 2005. 73: 2367–2378. [Бесплатная статья PMC] [PubMed] [Google Scholar] 76. Юн Дж. В., Миннич С. А., Ан Дж. С., Пак Ю. Х., Пащинский А., Ховде С. Дж.. Терморегуляция оперона Escherichia coli O157: H7 pO157 ecf и активность миристоилтрансферазы липида А включает в себя изогнутую по своей природе ДНК. Mol Microbiol. 2004. 51: 419–435. [PubMed] [Google Scholar] 77. Юк Х.Г., Маршалл Д.Л. Адаптация Escherichia coli O157: H7 к pH изменяет липидный состав мембран, секрецию веротоксина и устойчивость к симулированной кислоте желудочного сока.Appl Environ Microbiol. 2004. 70: 3500–3505. [Бесплатная статья PMC] [PubMed] [Google Scholar]
А., Ан Дж. С., Пак Ю. Х., Пащинский А., Ховде С. Дж.. Терморегуляция оперона Escherichia coli O157: H7 pO157 ecf и активность миристоилтрансферазы липида А включает в себя изогнутую по своей природе ДНК. Mol Microbiol. 2004. 51: 419–435. [PubMed] [Google Scholar] 77. Юк Х.Г., Маршалл Д.Л. Адаптация Escherichia coli O157: H7 к pH изменяет липидный состав мембран, секрецию веротоксина и устойчивость к симулированной кислоте желудочного сока.Appl Environ Microbiol. 2004. 70: 3500–3505. [Бесплатная статья PMC] [PubMed] [Google Scholar]E. coli
Обзор
Escherichia coli ( E. coli ) — это бактерия, которая обычно встречается в кишечнике людей и теплокровных животных. Большинство штаммов E. coli безвредны. Однако некоторые штаммы, такие как продуцирующая токсин шига E. coli (STEC), могут вызывать тяжелые болезни пищевого происхождения. Он передается человеку в первую очередь через употребление зараженных пищевых продуктов, таких как сырые или недоваренные мясные продукты, сырое молоко и зараженные сырые овощи и проростки.
STEC производит токсины, известные как шига-токсины из-за их сходства с токсинами, вырабатываемыми Shigella dysenteriae. STEC может расти в диапазоне температур от 7 ° C до 50 ° C, при оптимальной температуре 37 ° C. Некоторые STEC могут расти в кислых продуктах питания до pH 4,4 и в продуктах с минимальной активностью воды ( W ) 0,95.
STEC разрушается при тщательном приготовлении пищи до тех пор, пока все части не достигнут температуры 70 ° C или выше. E. coli O157: H7 — наиболее важный серотип STEC с точки зрения общественного здравоохранения; однако другие серотипы часто участвовали в спорадических случаях и вспышках.
Симптомы
Симптомы заболеваний, вызванных STEC, включают спазмы в животе и диарею, которые в некоторых случаях могут переходить в кровавую диарею (геморрагический колит). Также могут возникнуть жар и рвота. Инкубационный период может составлять от 3 до 8 дней, в среднем от 3 до 4 дней. Большинство пациентов выздоравливают в течение 10 дней, но у небольшой части пациентов (особенно у маленьких детей и пожилых людей) инфекция может привести к опасному для жизни заболеванию, например, гемолитико-уремическому синдрому (ГУС). ГУС характеризуется острой почечной недостаточностью, гемолитической анемией и тромбоцитопенией (низкий уровень тромбоцитов).
ГУС характеризуется острой почечной недостаточностью, гемолитической анемией и тромбоцитопенией (низкий уровень тромбоцитов).
Подсчитано, что до 10% пациентов с инфекцией STEC может развить ГУС, а уровень летальности составляет от 3 до 5%. В целом ГУС является наиболее частой причиной острой почечной недостаточности у детей раннего возраста. Это может вызвать неврологические осложнения (такие как судороги, инсульт и кома) у 25% пациентов с ГУС и хронические почечные осложнения, обычно легкие, примерно у 50% выживших.
Людям, у которых наблюдается кровавая диарея или сильные спазмы в животе, следует обратиться за медицинской помощью.Антибиотики не являются частью лечения пациентов с заболеванием STEC и могут увеличить риск последующего HUS.
Источники и передача
Большая часть доступной информации о STEC относится к серотипу O157: H7, поскольку биохимически он легко дифференцируется от других штаммов E. coli . Резервуаром этого патогена является в основном крупный рогатый скот. Кроме того, другие жвачные животные, такие как овцы, козы, олени, считаются значительными резервуарами, в то время как другие млекопитающие (например, свиньи, лошади, кролики, собаки и кошки) и птицы (например, куры и индейки) были обнаружены инфицированными.
Кроме того, другие жвачные животные, такие как овцы, козы, олени, считаются значительными резервуарами, в то время как другие млекопитающие (например, свиньи, лошади, кролики, собаки и кошки) и птицы (например, куры и индейки) были обнаружены инфицированными.
E. coli O157: H7 передается человеку в основном через употребление зараженных пищевых продуктов, таких как сырые или недоваренные мясные продукты и сырое молоко. Фекальное загрязнение воды и других пищевых продуктов, а также перекрестное заражение во время приготовления пищи (говядиной и другими мясными продуктами, загрязненными поверхностями и кухонной утварью) также могут привести к заражению. Примеры пищевых продуктов, вызывающих вспышки E. coli O157: H7, включают недоваренные гамбургеры, сушеную салями, непастеризованный яблочный сидр свежего отжима, йогурт и сыр из сырого молока.
Растущее число вспышек связано с потреблением фруктов и овощей (включая проростки, шпинат, салат, капустный салат и салат), причем заражение может быть связано с контактом с фекалиями домашних или диких животных на определенном этапе во время выращивания или обработки. STEC также был изолирован из водоемов (таких как пруды и ручьи), колодцев и желобов, и было обнаружено, что он выживает в течение нескольких месяцев в навозе и отложениях в водоемах. Сообщалось о передаче через воду как из загрязненной питьевой воды, так и из рекреационных вод.
STEC также был изолирован из водоемов (таких как пруды и ручьи), колодцев и желобов, и было обнаружено, что он выживает в течение нескольких месяцев в навозе и отложениях в водоемах. Сообщалось о передаче через воду как из загрязненной питьевой воды, так и из рекреационных вод.
Контакт между людьми — важный путь передачи орально-фекальным путем. Сообщалось о бессимптомном состоянии носительства, когда люди не проявляют клинических признаков заболевания, но способны инфицировать других. Продолжительность выведения STEC составляет около 1 недели или меньше у взрослых, но может быть дольше у детей. Посещение ферм и других мест, где население может напрямую контактировать с сельскохозяйственными животными, также было определено как важный фактор риска заражения STEC.
Профилактика
Профилактика инфекции требует мер контроля на всех этапах пищевой цепи, от сельскохозяйственного производства на ферме до обработки, производства и приготовления пищевых продуктов как на коммерческих предприятиях, так и на домашних кухнях.
Промышленность
Число случаев заболевания можно снизить с помощью различных стратегий смягчения последствий для говяжьего фарша (например, скрининга животных перед убоем для уменьшения попадания большого количества патогенов в среду убоя).Надлежащая гигиеническая практика забоя снижает загрязнение туш фекалиями, но не гарантирует отсутствие STEC в продуктах.
Для сведения микробиологического заражения к минимуму необходимо просвещение рабочих на фермах, скотобойнях и тех, кто занимается производством продуктов питания, по вопросам гигиенического обращения с пищевыми продуктами. Единственный эффективный метод удаления STEC из пищевых продуктов — это введение бактерицидной обработки, такой как нагревание (например, приготовление пищи или пастеризация) или облучение.
Дом
Профилактические меры против инфекции E. coli O157: H7 аналогичны мерам, рекомендованным для других болезней пищевого происхождения. Базовая практика надлежащей гигиены пищевых продуктов, описанная в документе ВОЗ « Пять ключей к более безопасным продуктам питания », может предотвратить передачу патогенов, ответственных за многие болезни пищевого происхождения, а также защитить от болезней пищевого происхождения, вызываемых STEC.
Пять ключей к безопасному питанию:
- Содержать в чистоте.
- Разделить сырые и приготовленные.
- Тщательно готовьте.
- Храните пищу при безопасной температуре.
- Используйте безопасную воду и сырье.
Такие рекомендации следует выполнять во всех случаях, особенно «тщательно готовить», чтобы температура в центре блюда была не менее 70 ° C. Обязательно тщательно мойте фрукты и овощи, особенно если их едят в сыром виде. По возможности овощи и фрукты следует очистить от кожуры. Уязвимые группы населения (например, маленькие дети и пожилые люди) должны избегать употребления сырых или недоваренных мясных продуктов, сырого молока и продуктов из сырого молока.
Настоятельно рекомендуется регулярное мытье рук, особенно перед приготовлением пищи или употреблением в пищу, а также после контакта с туалетом, особенно для людей, которые заботятся о маленьких детях, пожилых людях или лицах с ослабленным иммунитетом, поскольку бактерия может передаваться от человека к человеку, а также через еда, вода и прямой контакт с животными.
Ряд инфекций STEC был вызван контактом с рекреационной водой. Поэтому также важно защитить такие водные территории, а также источники питьевой воды от отходов животноводства (4).
Производители фруктов и овощей
Программа ВОЗ « Пять ключей к выращиванию более безопасных фруктов и овощей » предоставляет сельским работникам, выращивающим свежие фрукты и овощи для себя, своих семей и для продажи на местных рынках, основные методы предотвращения микробного заражения свежих продуктов во время посадки, выращивания и сбора урожая. и хранение.
Пять ключей к выращиванию более безопасных фруктов и овощей:
- Соблюдайте личную гигиену.
- Защищать поля от заражения фекалиями животных.
- Используйте обработанные фекальные отходы.
- Оценка и управление рисками, связанными с оросительной водой.
- Содержите инвентарь для сбора урожая и хранения в чистоте и сухости.
Ответ ВОЗ
ВОЗ предоставляет научные оценки для контроля STEC в пищевых продуктах. Эти оценки служат основой для международных пищевых стандартов, руководств и рекомендаций, разработанных Комиссией Codex Alimentarius.
Эти оценки служат основой для международных пищевых стандартов, руководств и рекомендаций, разработанных Комиссией Codex Alimentarius.
ВОЗ способствует укреплению систем безопасности пищевых продуктов, продвигая передовые методы производства и обучая розничных продавцов и потребителей правильному обращению с пищевыми продуктами и предотвращению загрязнения.
Во время вспышек E. coli , например, в Европе в 2011 году, ВОЗ поддерживает координацию обмена информацией и сотрудничества через Международные медико-санитарные правила и Международную сеть органов по безопасности пищевых продуктов (INFOSAN) во всем мире. ВОЗ тесно сотрудничает с национальными органами здравоохранения и международными партнерами, предоставляя техническую помощь и самую последнюю информацию о вспышках.
| Распечатать эту страницу
|


 coli (страница 1)
coli (страница 1) coli был показано, что это причина вспышки диареи среди младенцев.
coli был показано, что это причина вспышки диареи среди младенцев.  В полная последовательность оснований ДНК E.геном coli известен поскольку 1997 г.
В полная последовательность оснований ДНК E.геном coli известен поскольку 1997 г.
 Частично это адаптирует E. coli к кишечной (анаэробной) и это внекишечные (аэробные или анаэробные) среды обитания.
Частично это адаптирует E. coli к кишечной (анаэробной) и это внекишечные (аэробные или анаэробные) среды обитания. С его сложными механизмами для регулирование метаболизма бактерия может исследовать химический состав в своем среда перед синтезом любых ферментов, метаболизирующих эти соединения.Он не производит расточительно ферментов для разложения углерода. источники если они не доступны, и он не производит ферменты для синтез метаболитов, если они доступны в окружающей среде в качестве питательных веществ.
С его сложными механизмами для регулирование метаболизма бактерия может исследовать химический состав в своем среда перед синтезом любых ферментов, метаболизирующих эти соединения.Он не производит расточительно ферментов для разложения углерода. источники если они не доступны, и он не производит ферменты для синтез метаболитов, если они доступны в окружающей среде в качестве питательных веществ. coli в в кишечник и кал человека привели к отслеживанию бактерий в природе в качестве индикатор фекального загрязнения и загрязнения воды.Таким образом, это означает, что везде, где обнаруживается E. coli , может быть фекальный заражение кишечными паразитами человека.
coli в в кишечник и кал человека привели к отслеживанию бактерий в природе в качестве индикатор фекального загрязнения и загрязнения воды.Таким образом, это означает, что везде, где обнаруживается E. coli , может быть фекальный заражение кишечными паразитами человека.Типичная и атипичная энтеропатогенная Escherichia coli — Том 8, номер 5 — май 2002 г.
 — журнал Emerging Infectious Diseases
— журнал Emerging Infectious DiseasesПринадлежность к авторам: * Специальная лаборатория микробиологии, Институт Бутантана, Сан-Паулу, Бразилия; † Федеральный университет Сан-Паулу, Сан-Паулу, Бразилия;
Рисунок 1
Рисунок 1.Присоединение и стирание поражения, показывающее стирание микроворсинок (mv) и пьедестала (звездочка) с прилипшими энтеропатогенными бактериями Escherichia coli (EPEC) (стрелка). Перепечатано по ссылке 2 с разрешения директора …
Энтеропатогенный Escherichia coli (EPEC) является основной причиной детской диареи в развивающихся странах. В промышленно развитых странах частота этих организмов снизилась, но они продолжают оставаться важной причиной диареи ( 1 ).Центральным механизмом патогенеза EPEC является поражение, называемое прикреплением и стиранием (A / E), которое характеризуется разрушением микроворсинок, плотным прикреплением бактерий к кишечному эпителию, формированием пьедестала и агрегацией поляризованного актина и других элементов цитоскелета в места прикрепления бактерий (рисунок 1). Тест на флуоресцентное окрашивание актина позволяет идентифицировать штаммы, которые вызывают поражения A / E, путем обнаружения агрегированных актиновых филаментов под прикрепленными бактериями ( 3 ).Способность образовывать A / E-поражения также была обнаружена у штаммов штаммов E. coli , продуцирующих токсин Shiga (энтерогеморрагическая E. coli [EHEC]), и у штаммов других видов бактерий (1).
Тест на флуоресцентное окрашивание актина позволяет идентифицировать штаммы, которые вызывают поражения A / E, путем обнаружения агрегированных актиновых филаментов под прикрепленными бактериями ( 3 ).Способность образовывать A / E-поражения также была обнаружена у штаммов штаммов E. coli , продуцирующих токсин Shiga (энтерогеморрагическая E. coli [EHEC]), и у штаммов других видов бактерий (1).
Рисунок 2
Рисунок 2. Схема основных генов локуса области удаления энтероцитов (LEE) и энтеропатогенной плазмиды фактора адгезии (EAF) Escherichia coli (EPEC).
Генетические детерминанты образования поражений A / E расположены в локусе сглаживания энтероцитов (LEE) ( 4 ), острове патогенности, который содержит гены, кодирующие интимин, систему секреции типа III, ряд секретируемых (Esp) и транслоцированный рецептор интимина под названием Tir ( 1 ) (рис. 2).Два сайта вставки LEE были описаны на хромосоме E. coli , и сообщалось о третьем неидентифицированном сайте вставки ( 5 ).
Интимин, белок внешней мембраны массой 94 кДа, кодируемый геном eae , отвечает за плотное прилегание бактерий к мембранам энтероцитов. Изучение антигенных вариаций в 280-аминокислотных остатках С-концевой части интимина (рецептор-связывающего домена белка) и использование анализа полимеразной цепной реакции позволяет классифицировать различные типы или подтипы интимина среди EPEC и STEC. штаммы ( 6 ).Молекулы Esp (EspA, B и D) участвуют в образовании транслокона, который доставляет эффекторные молекулы к клетке-хозяину и разрушает цитоскелет, нарушая функции клетки-хозяина ( 7 ). Tir, который является одним из белков, перемещаемых EPEC, внедряется в мембрану клетки-хозяина, где он действует как рецептор к интимину ( 8 ).
Многие штаммы EPEC образуют характерный образец адгезии, называемый локализованной адгезией, в клетках культуры ткани ( 9 ).По этой схеме бактерии связываются с локализованными участками поверхности клетки, образуя компактные микроколонии (бактериальные кластеры), которые можно визуализировать после контакта бактерий с клетками в течение 3 часов. Этот феномен связан с присутствием плазмиды большого фактора адгезии EPEC (EAF), которая несет так называемую последовательность EAF (рисунок 2) ( 1 ). В плазмиде EAF также присутствует кластер генов, которые кодируют образующие пучки пили (BFP), которые связывают бактерии внутри микроколоний и, таким образом, способствуют их стабилизации ( 1 ).
Этот феномен связан с присутствием плазмиды большого фактора адгезии EPEC (EAF), которая несет так называемую последовательность EAF (рисунок 2) ( 1 ). В плазмиде EAF также присутствует кластер генов, которые кодируют образующие пучки пили (BFP), которые связывают бактерии внутри микроколоний и, таким образом, способствуют их стабилизации ( 1 ).
Плазмида EAF не важна для образования повреждений A / E, хотя ее присутствие увеличивает их эффективность, вероятно, за счет влияния кластера плазмидных регуляторных генов ( на A , B , C ), которые увеличивают экспрессию хромосомных генов LEE ( 1 ). Доказательства также указывают на то, что BFP играет роль в адгезии клеток-хозяев, которая аналогичным образом увеличивает эффективность образования повреждений A / E ( 7 ).
В 1995 году во время Второго международного симпозиума по EPEC в Сан-Паулу большинство участников приняли следующее определение EPEC: «EPEC являются диареогенными Escherichia coli , которые вызывают характерную гистопатологию, известную как прикрепление и стирание (A / E) на клетках кишечника и которые не производят шига, шига-подобные или вероцитотоксины. Типичный ЕРЕС человеческого происхождения обладает плазмидой вирулентности, известной как плазмида EAF (фактор адгезии ЕРЕС), которая кодирует локализованную адгезию к культивируемым эпителиальным клеткам, опосредованную.. . BFP, в то время как атипичный EPEC не обладает этой плазмидой. Большинство типичных штаммов EPEC попадают в некоторые хорошо известные серотипы O: H »( 10 ). Согласно этому определению, основное различие между типичным и атипичным EPEC заключается в наличии плазмиды EAF в первой группе организмов и ее отсутствии во второй.
Типичный ЕРЕС человеческого происхождения обладает плазмидой вирулентности, известной как плазмида EAF (фактор адгезии ЕРЕС), которая кодирует локализованную адгезию к культивируемым эпителиальным клеткам, опосредованную.. . BFP, в то время как атипичный EPEC не обладает этой плазмидой. Большинство типичных штаммов EPEC попадают в некоторые хорошо известные серотипы O: H »( 10 ). Согласно этому определению, основное различие между типичным и атипичным EPEC заключается в наличии плазмиды EAF в первой группе организмов и ее отсутствии во второй.
Наиболее изученные штаммы EPEC принадлежат к серии O антигенных групп, известных как серогруппы EPEC O. Двенадцать серогрупп EPEC были признаны Всемирной организацией здравоохранения в 1987 году: O26, O55, O86, O111, O114, O119, O125, O126, O127, O128, O142 и O158.Эти серогруппы включают как типичные, так и атипичные штаммы EPEC, а также другие диареогенные категории E. coli , в основном энтероагрегантные E. coli (EAEC) ( 11 — 14 ). Кроме того, большинство штаммов каждой категории соответствуют определенным серотипам в каждой серогруппе O. Разделение штаммов EPEC на типичные и атипичные имеет важные последствия, которые еще не полностью оценены. EPEC больше нельзя рассматривать как единую группу энтеропатогенных организмов.Целью данной статьи является обзор основных различий между типичным и атипичным EPEC, которые следует учитывать при исследованиях с участием этих организмов.
Кроме того, большинство штаммов каждой категории соответствуют определенным серотипам в каждой серогруппе O. Разделение штаммов EPEC на типичные и атипичные имеет важные последствия, которые еще не полностью оценены. EPEC больше нельзя рассматривать как единую группу энтеропатогенных организмов.Целью данной статьи является обзор основных различий между типичным и атипичным EPEC, которые следует учитывать при исследованиях с участием этих организмов.
Типичные и атипичные штаммы EPEC относятся к двум разным наборам серотипов (Таблица 1). Эта таблица была построена на основе аналогичных исследований, проведенных в Сан-Паулу ( 11 — 15 ) и Великобритании ( 14 ) и в меньшем масштабе в Рио-де-Жанейро ( 16 ) и Италии ( 17 ).Большинство типичных штаммов было выделено в Сан-Паулу и Рио-де-Жанейро, а большинство нетипичных — в Великобритании и Италии. Серотипы, выделенные в Сан-Паулу, включают подвижные и неподвижные штаммы (обозначенные помещением антигена H в скобки). Антигены H этих неподвижных штаммов были выведены с помощью рестрикционного анализа генов fliC (B.A. Botelho, et al., Неопубликованные данные). Эти серотипы могут включать как подвижные, так и неподвижные варианты (таблица 1).
Антигены H этих неподвижных штаммов были выведены с помощью рестрикционного анализа генов fliC (B.A. Botelho, et al., Неопубликованные данные). Эти серотипы могут включать как подвижные, так и неподвижные варианты (таблица 1).
Большинство серотипов в таблице 1 можно легко классифицировать как типичные или атипичные.Однако некоторые серотипы не так легко классифицировать, в основном те, которые включают штаммы, продуцирующие Stx, из которых наиболее частыми являются серотипы O26: H- и h21 и O111ac: [H8] (рассматриваемые некоторыми авторами как EHEC или STEC) ( 1 ). Фактически, эти и другие серотипы со свойствами, аналогичными свойствам O128: h3, не являются истинными атипичными серотипами EPEC или STEC, а скорее являются гетерогенными серотипами, которые включают различные клоны или генетические линии. Например, мы недавно показали с помощью случайной амплифицированной полиморфной ДНК, что штаммы, продуцирующие O26: h21 Stx, выделенные в Европе и Северной Америке, генетически отличаются от Stx-отрицательных штаммов того же серотипа, выделенных в Бразилии ( 18 ).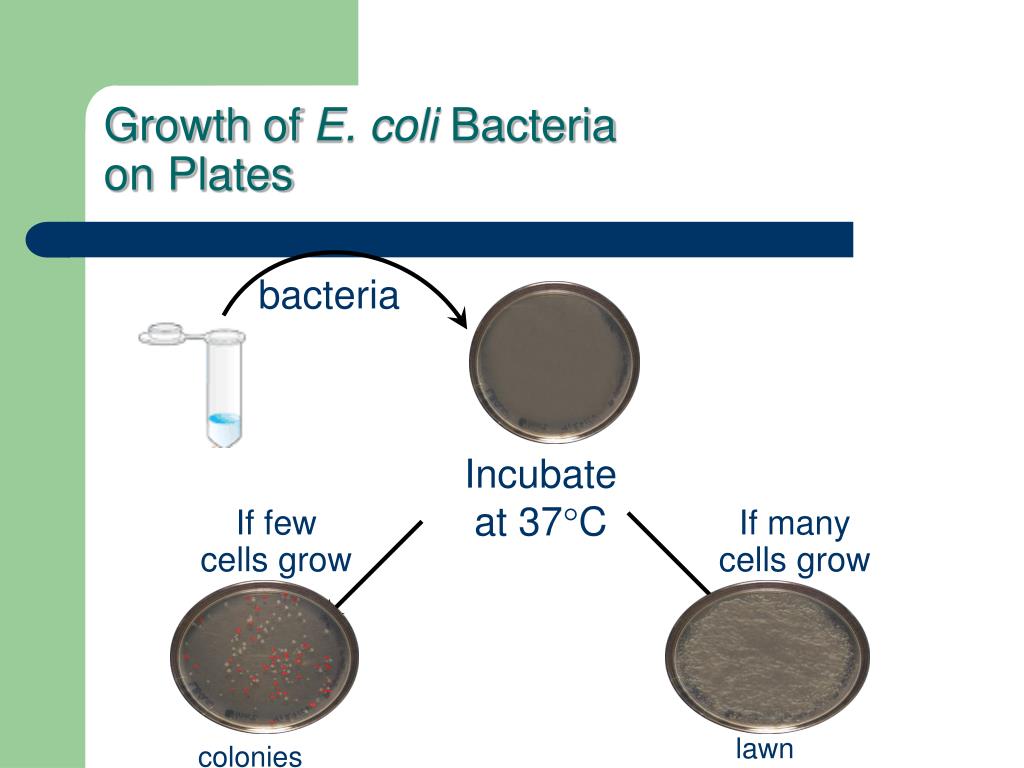 Хотя такого рода исследования не проводились с серотипом O128: h3, этот серотип также является гетерогенным, поскольку он включает различные риботипы с различными характеристиками вирулентности (L.R. Trabulsi et al., Неопубликованные данные). Некоторые клоны, продуцирующие Stx, имеют нерегулярное географическое распространение, поэтому их можно найти в некоторых странах, но не в других. Другие характеристики, которые могут затруднить различение типичного EPEC от атипичного, связаны с маркерами плазмиды EAF. Например, серотипы O119: h3 и O128: h3 реагируют с зондом bfpA , но не имеют настоящей плазмиды EAF.Эти серотипы имеют плазмиду 100-MDa, которая не содержит оперона bfp и, следовательно, не продуцирует BFP ( 19 ). Напротив, некоторые штаммы O142: H6 не реагируют с зондом EAF, но продуцируют BFP и демонстрируют типичный образец локализованного прилипания (LA). Эти штаммы могут иметь плазмиду EAF с дефектом в области EAF, которая не мешает функциям плазмиды.
Хотя такого рода исследования не проводились с серотипом O128: h3, этот серотип также является гетерогенным, поскольку он включает различные риботипы с различными характеристиками вирулентности (L.R. Trabulsi et al., Неопубликованные данные). Некоторые клоны, продуцирующие Stx, имеют нерегулярное географическое распространение, поэтому их можно найти в некоторых странах, но не в других. Другие характеристики, которые могут затруднить различение типичного EPEC от атипичного, связаны с маркерами плазмиды EAF. Например, серотипы O119: h3 и O128: h3 реагируют с зондом bfpA , но не имеют настоящей плазмиды EAF.Эти серотипы имеют плазмиду 100-MDa, которая не содержит оперона bfp и, следовательно, не продуцирует BFP ( 19 ). Напротив, некоторые штаммы O142: H6 не реагируют с зондом EAF, но продуцируют BFP и демонстрируют типичный образец локализованного прилипания (LA). Эти штаммы могут иметь плазмиду EAF с дефектом в области EAF, которая не мешает функциям плазмиды. Возможно, лучшей отличительной характеристикой для типичных и атипичных серотипов EPEC будет продукция или отсутствие продукции BFP.
Возможно, лучшей отличительной характеристикой для типичных и атипичных серотипов EPEC будет продукция или отсутствие продукции BFP.
Характеристики вирулентности
В целом, типичные штаммы EPEC более однородны по своим характеристикам вирулентности, чем атипичные. За некоторыми исключениями, типичные штаммы продуцируют только факторы вирулентности, кодируемые областью LEE и плазмидой EAF. Исключения составляют продукция цитотоксического растягивающего токсина (CDT) всеми штаммами O86: h44 (LR Trabulsi et al., Неопубликованные данные) и продукция энтероагрегативного термостабильного токсина (EAST1) некоторыми штаммами серотипов O55: H6. и O127: H6 (T.А. Т. Гомес и др., Неопубликован. data), которые являются потенциальными факторами вирулентности. Напротив, атипичные штаммы EPEC часто экспрессируют EAST1 и другие потенциальные факторы вирулентности, не кодируемые в области LEE (Таблица 2). Соответственно, существует два типа атипичных штаммов EPEC: те, которые экспрессируют только факторы вирулентности, кодируемые LEE, и те, которые экспрессируют как LEE, так и факторы вирулентности, не кодируемые LEE. Обычно оба вида штаммов относятся к одному клону ( 11 , 12 , 15 ).Все атипичные серотипы EPEC, за исключением O125ac: H6, включают оба вида штаммов. Все штаммы этого серотипа, исследованные до сих пор, демонстрируют агрегированный характер приверженности и область LEE. Еще одним интересным различием между типичным и атипичным EPEC является наличие более чем одного вида штаммов в большинстве атипичных серотипов.
Обычно оба вида штаммов относятся к одному клону ( 11 , 12 , 15 ).Все атипичные серотипы EPEC, за исключением O125ac: H6, включают оба вида штаммов. Все штаммы этого серотипа, исследованные до сих пор, демонстрируют агрегированный характер приверженности и область LEE. Еще одним интересным различием между типичным и атипичным EPEC является наличие более чем одного вида штаммов в большинстве атипичных серотипов.
Рисунок 3
Рисунок 3. Характер адгезии энтеропатогенных штаммов Escherichia coli (EPEC). Локализованная приверженность (LA), диффузная приверженность (DA), агрегативная приверженность (AA) и подобная локализованной приверженности (LAL).Увеличение: X100.
Типичные и атипичные штаммы EPEC также различаются по характеру адгезии. Типичные штаммы демонстрируют только паттерн LA, в то время как атипичные штаммы могут демонстрировать паттерн LAL (локализованное прилипание) ( 12 ), паттерн DA (диффузное прилипание) или паттерн AA (агрегативное прилипание) (рисунок 3). Паттерн КЛЛ характерен для штаммов большинства серотипов и опосредуется в основном интимином ( 20 ). Модель DA опосредуется адгезином Afa (R.Keller et al., Unpub. data), а AA опосредуется агрегированным адгезином. Ген cdt серотипа O86: h44 (LR Trabulsi et al., Unpub. Data) и afa ген серотипа O55: H7 расположены на бактериальной хромосоме (R. Keller et al., Unpub. Data) . Типичные и атипичные EPEC также имеют некоторые интересные различия в отношении типов интимина (таблица 3).
Паттерн КЛЛ характерен для штаммов большинства серотипов и опосредуется в основном интимином ( 20 ). Модель DA опосредуется адгезином Afa (R.Keller et al., Unpub. data), а AA опосредуется агрегированным адгезином. Ген cdt серотипа O86: h44 (LR Trabulsi et al., Unpub. Data) и afa ген серотипа O55: H7 расположены на бактериальной хромосоме (R. Keller et al., Unpub. Data) . Типичные и атипичные EPEC также имеют некоторые интересные различия в отношении типов интимина (таблица 3).
Рисунок 4
Рисунок 4. Дендограмма, иллюстрирующая генетические различия между типичными и атипичными энтеропатогенными штаммами Escherichia coli (EPEC) и E.coli O157: штаммы H7.
Чтобы исследовать генетические отношения между типичными и атипичными штаммами EPEC, мы использовали случайную амплифицированную полиморфную ДНК для изучения нашей коллекции штаммов, которая включает большинство серотипов, показанных в таблице 1. Дендрограмма, полученная на основе этих данных (Рисунок 4), показывает, что наиболее типичные и атипичные штаммы принадлежат к разным генетическим группам, и что атипичные штаммы ближе к штаммам серотипа O157: H7 (STEC), которые были включены в исследование в целях сравнения (S. Y.Bando et al., Неопуб. данные). Единственными исключениями были типичные и атипичные штаммы h3, которые не разделялись и составляли подгруппу в группе атипичных / STEC. Общие результаты этого исследования напоминают результаты, сообщенные Whittam et al. ( 21 ), который использовал мультилокусный ферментный электрофорез для изучения аналогичной коллекции штаммов и выделил четыре генетические группы: EPEC 1 (штаммы H6 / h44), EPEC 2 (штаммы h3), EHEC 1 (O55: H7 и O157: H7). штаммы) и EHEC 2 (штаммы O26: h21 и O111ac: H-).Группа EPEC 2 также была ближе к группам EHEC. В этой статье мы не использовали разделение EPEC на EPEC 1 и EPEC 2, но это может быть важно в будущем. Между двумя клональными группами существует несколько других различий (R. Keller et al., Неопубликованные данные). Что касается эпидемиологии, серотип EPEC 2 (O111: h3) сильно связан с внутрибольничной инфекцией, в то время как серотип EPEC 1 (O119: H6) сильнее связан с инфекцией в сообществе ( 22 ).
Y.Bando et al., Неопуб. данные). Единственными исключениями были типичные и атипичные штаммы h3, которые не разделялись и составляли подгруппу в группе атипичных / STEC. Общие результаты этого исследования напоминают результаты, сообщенные Whittam et al. ( 21 ), который использовал мультилокусный ферментный электрофорез для изучения аналогичной коллекции штаммов и выделил четыре генетические группы: EPEC 1 (штаммы H6 / h44), EPEC 2 (штаммы h3), EHEC 1 (O55: H7 и O157: H7). штаммы) и EHEC 2 (штаммы O26: h21 и O111ac: H-).Группа EPEC 2 также была ближе к группам EHEC. В этой статье мы не использовали разделение EPEC на EPEC 1 и EPEC 2, но это может быть важно в будущем. Между двумя клональными группами существует несколько других различий (R. Keller et al., Неопубликованные данные). Что касается эпидемиологии, серотип EPEC 2 (O111: h3) сильно связан с внутрибольничной инфекцией, в то время как серотип EPEC 1 (O119: H6) сильнее связан с инфекцией в сообществе ( 22 ).
Патогенность большинства типичных серотипов EPEC подтверждена волонтерскими исследованиями ( 1 ). Что касается атипичного EPEC, нам известно только об одном исследовании с участием добровольцев, которое было проведено Levine et al. ( 23 ) со штаммом O128: h3. Этот штамм вводили в разных дозах 15 взрослым добровольцам, ни один из которых не заболел. Хотя это исследование было тщательно проведено, его результаты трудно оценить, поскольку характеристики вирулентности штамма не были известны, а серотип O128: h3 может включать невирулентные штаммы ( 24 ).
Что касается атипичного EPEC, нам известно только об одном исследовании с участием добровольцев, которое было проведено Levine et al. ( 23 ) со штаммом O128: h3. Этот штамм вводили в разных дозах 15 взрослым добровольцам, ни один из которых не заболел. Хотя это исследование было тщательно проведено, его результаты трудно оценить, поскольку характеристики вирулентности штамма не были известны, а серотип O128: h3 может включать невирулентные штаммы ( 24 ).
Атипичные штаммы EPEC могут быть менее вирулентными, чем типичные.Одной из причин может быть отсутствие плазмиды EAF; Левин и др. ( 25 ) показали, что штамм O127: H6 без плазмиды был менее вирулентным для взрослых добровольцев, чем штамм дикого типа. Однако не было доказано, что атипичные штаммы EPEC менее патогенны, и у этих организмов есть другие факторы вирулентности, которые могут компенсировать отсутствие плазмиды EAF. Для решения этой проблемы необходимы дополнительные исследования.
Связь с диареей
Типичные серотипы EPEC тесно связаны с диареей у детей младше 1 года. В нескольких контролируемых исследованиях, проведенных в Бразилии, эти серотипы были признаны основной причиной эндемической диареи в этой возрастной группе ( 26 , 27 ). Частота типичных серотипов EPEC у детей старше 1 года ниже и аналогична частоте в контроле (2–4%). Инфекции взрослых редки и обычно связаны с другими состояниями ( 1 ). Повышенная резистентность у детей старшего возраста и взрослых может быть связана с развитием иммунитета или потерей рецепторов для некоторых специфических адгезинов ( 1 ).
В нескольких контролируемых исследованиях, проведенных в Бразилии, эти серотипы были признаны основной причиной эндемической диареи в этой возрастной группе ( 26 , 27 ). Частота типичных серотипов EPEC у детей старше 1 года ниже и аналогична частоте в контроле (2–4%). Инфекции взрослых редки и обычно связаны с другими состояниями ( 1 ). Повышенная резистентность у детей старшего возраста и взрослых может быть связана с развитием иммунитета или потерей рецепторов для некоторых специфических адгезинов ( 1 ).
Что касается иммунитета, то несколько исследований, проведенных в Бразилии ( 28 ) и совсем недавно в Мексике ( 29 ), показали, что у детей вырабатываются высокие уровни антител против основных факторов вирулентности EPEC. Кроме того, молозиво матерей, живущих в эндемичных районах, очень богато антителами иммуноглобулина А против факторов вирулентности EPEC ( 28 — 30 ). Гораздо меньше известно о связи атипичных серотипов с диареей, но обычно эти серотипы выделяют от детей с диареей, которые не являются носителями других энтеропатогенных агентов. Сильная связь атипичных серотипов EPEC с эндемической диареей еще не продемонстрирована. Однако в Финляндии была описана крупная вспышка диареи, вызванной серотипом O111: H9 ( 31 ).
Сильная связь атипичных серотипов EPEC с эндемической диареей еще не продемонстрирована. Однако в Финляндии была описана крупная вспышка диареи, вызванной серотипом O111: H9 ( 31 ).
Распространенность в развивающихся и промышленно развитых странах
Заметным эпидемиологическим различием между типичными и атипичными серотипами EPEC является их географическое распределение. Типичные серотипы EPEC традиционно были связаны со вспышками детской диареи, и, фактически, первые штаммы EPEC, выделенные в разных странах, были серотипами O55: H6 и O111: h3 ( 32 ).В прошлом эти эпидемические серотипы часто определялись в промышленно развитых странах как причина вспышек и спорадических случаев диареи, но в настоящее время они очень редки ( 1 ). В этих странах сегодня преобладают серотипы без плазмиды EAF ( 14 , 33 ). В Соединенном Королевстве, например, EAF-положительные штаммы составляют только 10% всех штаммов EPEC ( 14 ). Ситуация в развивающихся странах четко не определена, но несколько исследований в Бразилии в 1980-х и начале 1990-х годов показали высокую частоту типичных серотипов ( 34 , 35 ).Однако некоторые недавние исследования показали очень низкую частоту типичных EPEC и относительно высокую частоту атипичных EPEC (L.C. Campos, личные сообщения и неопубликованные данные). Этот результат совпадает с уменьшением числа случаев диареи в нескольких регионах Бразилии, что позволяет предположить, что изменения, произошедшие в промышленно развитых странах, вероятно, уже происходят в Бразилии. Причина этих изменений не ясна, но снижение частоты EAF-положительных серотипов, которое произошло в Европе и Соединенных Штатах и начало встречаться в Бразилии, может быть связано с улучшением терапии, санитарных условий и контроля. госпитальных инфекций.С другой стороны, появление и рост частоты атипичных штаммов EPEC может иметь происхождение, аналогичное тем, которые привели к появлению и увеличению частоты O157: H7 и других серотипов STEC ( 35 ).
Ситуация в развивающихся странах четко не определена, но несколько исследований в Бразилии в 1980-х и начале 1990-х годов показали высокую частоту типичных серотипов ( 34 , 35 ).Однако некоторые недавние исследования показали очень низкую частоту типичных EPEC и относительно высокую частоту атипичных EPEC (L.C. Campos, личные сообщения и неопубликованные данные). Этот результат совпадает с уменьшением числа случаев диареи в нескольких регионах Бразилии, что позволяет предположить, что изменения, произошедшие в промышленно развитых странах, вероятно, уже происходят в Бразилии. Причина этих изменений не ясна, но снижение частоты EAF-положительных серотипов, которое произошло в Европе и Соединенных Штатах и начало встречаться в Бразилии, может быть связано с улучшением терапии, санитарных условий и контроля. госпитальных инфекций.С другой стороны, появление и рост частоты атипичных штаммов EPEC может иметь происхождение, аналогичное тем, которые привели к появлению и увеличению частоты O157: H7 и других серотипов STEC ( 35 ).
Типичные серотипы EPEC не были обнаружены у животных ( 1 ), что позволяет предположить, что человек является единственным живым резервуаром для этих организмов. Напротив, большинство атипичных серотипов EPEC были выделены от разных видов животных. Связь между серотипом O26: h21 и телятами хорошо известна ( 36 ).Недавние исследования подчеркнули выделение штаммов, продуцирующих Stx, из-за их роли в гемолитико-уремическом синдроме, но от крупного рогатого скота было выделено eae -положительных, Stx-отрицательных штаммов ( 37 ). Этот вид штамма следует считать атипичным EPEC. Аналогичная ситуация существует в отношении серотипа O111ac, и 69 штаммов O111ac, о которых сообщают Ewing et al. в 1963 г. все были изолированы от обезьян ( 38 ).
Серотип O128: h3 довольно часто встречается у кроликов и собак и, как и штаммы человека, выделенные в Бразилии, является отрицательным по EAF (Pestana de Castro, pers.общ.). В недавнем исследовании группы Пестана де Кастро серотипы O119: h3 и O111: h35 (EAF-отрицательный серотип, редко встречающийся в Бразилии, но часто встречающийся в Соединенном Королевстве), часто выделялись у собак. Необходимы дополнительные исследования распространенности атипичных серотипов EPEC у животных, но имеющиеся данные убедительно свидетельствуют о том, что основным резервуаром для этих организмов являются разные виды животных, как в случае со штаммами STEC.
Необходимы дополнительные исследования распространенности атипичных серотипов EPEC у животных, но имеющиеся данные убедительно свидетельствуют о том, что основным резервуаром для этих организмов являются разные виды животных, как в случае со штаммами STEC.
Stx-Negative и
eae -Positive E. coli Штаммы в серогруппах, не относящихся к EPEC OИ stx-отрицательный, и eae -положительный E.coli обнаружены во многих серогруппах, не относящихся к EPEC O ( 39 ). Мы обнаружили такие штаммы более чем в 30 серогруппах E. coli O, и большая часть штаммов не агглютинировала в обычном наборе из антисывороток E. coli O. Некоторые штаммы реагируют с зондом EAF ( eae +, штаммы EAF +), но большинство не реагируют с этим зондом ( eae +, штаммы EAF-). За некоторыми исключениями, сообщалось только об одном или двух штаммах каждого из этих серотипов ( 40 ).
Дополнительные характеристики вирулентности штаммов eae +, EAF + не изучались, но недавно мы исследовали профиль вирулентности 49 различных штаммов eae +, EAF-, выделенных от детей с диареей в Сан-Паулу. Профиль был аналогичен профилю атипичного EPEC: многие штаммы были EAST1 + и E-hly +, а некоторые выражали паттерн приверженности либо AA, либо DA. Некоторые штаммы имели последовательность гамма-интимина, а у многих штаммов тип интимина не удалось идентифицировать.
Профиль был аналогичен профилю атипичного EPEC: многие штаммы были EAST1 + и E-hly +, а некоторые выражали паттерн приверженности либо AA, либо DA. Некоторые штаммы имели последовательность гамма-интимина, а у многих штаммов тип интимина не удалось идентифицировать.
Некоторые из этих штаммов действительно соответствуют типичным или атипичным EPEC, и необходимы дополнительные исследования для установления точной концепции для них, особенно для EAF-отрицательных штаммов. Некоторые из них, вероятно, являются штаммами STEC, утратившими генов stx ; мы не можем исключить возможность того, что штаммы DA и AA не являются истинными EAEC или DAEC, которые получили остров патогенности LEE путем горизонтального переноса. Для атипичных EPEC ситуация совершенно иная, поскольку было изучено большее количество штаммов, и большинство из них относятся к хорошо изученным серотипам.
Роль, которую играют эти штаммы EAF + и EAF- вне серогрупп EPEC, в эндемической диарее не установлена. В целом, штаммы редко выделяются из случаев диареи и из контрольной группы, и общая разница не является статистически значимой. Однако некоторые серотипов eae +, EAF +, а также несколько eae +, EAF- штаммов со специфическими профилями вирулентности, по-видимому, связаны с эндемической диареей ( 2 , 33 , 40 ). Что касается вспышек, то серотип eae +, EAF- (O39: H-) стал причиной вспышки диареи пищевого происхождения в 1991 г., в которой участвовало 100 взрослых в Миннесоте ( 41 ).
В целом, штаммы редко выделяются из случаев диареи и из контрольной группы, и общая разница не является статистически значимой. Однако некоторые серотипов eae +, EAF +, а также несколько eae +, EAF- штаммов со специфическими профилями вирулентности, по-видимому, связаны с эндемической диареей ( 2 , 33 , 40 ). Что касается вспышек, то серотип eae +, EAF- (O39: H-) стал причиной вспышки диареи пищевого происхождения в 1991 г., в которой участвовало 100 взрослых в Миннесоте ( 41 ).
Типичный и атипичный EPEC, по-видимому, составляют две группы отдельных организмов, которые имеют общий остров патогенности LEE. Атипичные EPEC ближе к STEC по генетическим характеристикам, серотипам, выработке токсинов, резервуару и другим эпидемиологическим аспектам. Как STEC, они напоминают появляющиеся патогены. В промышленно развитых странах они стали более частой причиной диареи, чем типичные EPEC, и такой же сдвиг может происходить в Бразилии. Большое количество Stx-отрицательных, eae -положительных типичных и атипичных EPEC-подобных штаммов за пределами серогрупп EPEC O, а также атипичных штаммов EPEC требуют дальнейшего изучения в отношении их вирулентности и эпидемиологического значения.
Большое количество Stx-отрицательных, eae -положительных типичных и атипичных EPEC-подобных штаммов за пределами серогрупп EPEC O, а также атипичных штаммов EPEC требуют дальнейшего изучения в отношении их вирулентности и эпидемиологического значения.
Доктор Трабульси — заслуженный профессор Университета Сан-Паулу и директор Лаборатории особой микробиологии до Института Бутантана, Сан-Паулу.
Вершина
Выводы, выводы и мнения, выраженные авторами, пишущими для этого журнала, не обязательно отражают официальную позицию Министерства здравоохранения и социальных служб США, Службы общественного здравоохранения, Центров по контролю и профилактике заболеваний или аффилированных с авторами учреждения.Торговые наименования используются только для идентификации и не подразумевают одобрения какой-либо из вышеперечисленных групп.
Характеристики диареи, вызывающей кишечную палочку, у детей младше 5 лет с острой диареей: исследование на базе больниц | BMC Infectious Diseases
 org/ScholarlyArticle»> 1.
org/ScholarlyArticle»> 1.Kotloff KL, Nataro JP, Blackwelder WC, Nasrin D, Farag TH, Panchalingam S, Wu Y, Sow SO, Sur D, Breiman RF, et al. Бремя и этиология диарейных заболеваний у младенцев и детей раннего возраста в развивающихся странах (глобальное многоцентровое исследование кишечника, GEMS): проспективное исследование случай-контроль.Ланцет. 2013. 382 (9888): 209–22.
Артикул PubMed Google ученый
Лю Л., Джонсон Х.Л., Кузенс С., Перин Дж., Скотт С., Лоун Дж. Э., Рудан И., Кэмпбелл Х., Сибулскис Р., Ли М. и др. Глобальные, региональные и национальные причины детской смертности: обновленный систематический анализ за 2010 год с указанием временных тенденций с 2000 года. Lancet (Лондон, Англия). 2012. 379 (9832): 2151–61.
Артикул Google ученый
 org/ScholarlyArticle»> 3.
org/ScholarlyArticle»> 3.Walker CL, Rudan I, Liu L, Nair H, Theodoratou E, Bhutta ZA, O’Brien KL, Campbell H, Black RE. Глобальное бремя детской пневмонии и диареи. Ланцет (Лондон, Англия). 2013. 381 (9875): 1405–16.
Артикул Google ученый
Yu J, Jing H, Lai S, Xu W., Li M, Wu J, Liu W., Yuan Z, Chen Y, Zhao S, et al. Этиология диареи у детей в возрасте до пяти лет в Китае: результаты пятилетнего наблюдения. J Infect.2015; 71 (1): 19–27.
Артикул PubMed PubMed Central Google ученый
Lanata CF, Fischer-Walker CL, Olascoaga AC, Torres CX, Aryee MJ, Black RE, Справочная группа по эпидемиологии здоровья детей Всемирного здравоохранения O, Unicef. Глобальные причины смертности от диарейных болезней у детей младше 5 лет: систематический обзор. PLoS One. 2013; 8 (9): e72788.
PLoS One. 2013; 8 (9): e72788.
CAS Статья PubMed PubMed Central Google ученый
Canizalez-Roman A, Flores-Villasenor HM, Gonzalez-Nunez E, Velazquez-Roman J, Vidal JE, Muro-Amador S, Alapizco-Castro G, Diaz-Quinonez JA, Leon-Sicairos N. Наблюдение за диарейными штаммами Escherichia Coli изолирован от случаев диареи у детей, взрослых и пожилых людей на северо-западе Мексики. Front Microbiol. 2016; 7: 1924.
Артикул PubMed PubMed Central Google ученый
Чжан И, Чжао И, Дин К., Ван Х, Чен Х, Лю И, Чен Ю.Анализ бактериальных патогенов, вызывающих острую диарею, на основе дозорного эпиднадзора в Шанхае, Китай, 2006-2011 гг. Jpn J Infect Dis. 2014; 67 (4): 264–8.
Артикул PubMed Google ученый
Джафари Ф., Гарсия-Гил Л. Дж., Салманзаде-Ахраби С., Шокрзаде Л., Аслани М. М., Поурхосинголи М. А., Дерахшан Ф., Зали М. Р.. Диагностика и распространенность энтеропатогенных бактерий у детей младше 5 лет с острой диареей в детских больницах Тегерана.J Infect. 2009. 58 (1): 21–7.
CAS Статья PubMed Google ученый
Саид А., Абд Х., Сандстром Г. Микробная этиология острой диареи у детей в возрасте до пяти лет в Хартуме, Судан. J Med Microbiol. 2015; 64 (Pt 4): 432–7.
Артикул PubMed PubMed Central Google ученый
 org/ScholarlyArticle»> 10.
org/ScholarlyArticle»> 10.Кроксен М.А., Лоу Р.Дж., Шольц Р., Кини К.М., Влодарска М., Финли ББ.Последние достижения в понимании кишечной патогенной кишечной палочки. Clin Microbiol Rev.2013; 26 (4): 822–80.
CAS Статья PubMed PubMed Central Google ученый
Zheng S, Yu F, Chen X, Cui D, Cheng Y, Xie G, Yang X, Han D, Wang Y, Zhang W. и др. Энтеропатогены у детей младше 5 лет с острой диареей: 5-летнее эпиднадзорное исследование на юго-восточном побережье Китая. BMC Infect Dis. 2016; 16 (1): 434.
Артикул PubMed PubMed Central Google ученый
Диас Р.К., Дос-Сантос, Британская Колумбия, Душ-Сантос Л.Ф., Виейра М.А., Яматоги Р.С., Монделли А.Л., Садацунэ Т. , Сфорчин Дж.М., Гомеш Т.А., Эрнандес РТ. Исследование патотипов диареи, вызывающей кишечную палочку, выявило атипичные энтеропатогенные кишечные палочки как предполагаемые возбудители диареи у детей, живущих в Ботукату, штат Сан-Паулу, Бразилия. АПМИС. 2016; 124 (4): 299–308.
, Сфорчин Дж.М., Гомеш Т.А., Эрнандес РТ. Исследование патотипов диареи, вызывающей кишечную палочку, выявило атипичные энтеропатогенные кишечные палочки как предполагаемые возбудители диареи у детей, живущих в Ботукату, штат Сан-Паулу, Бразилия. АПМИС. 2016; 124 (4): 299–308.
CAS Статья PubMed Google ученый
Гомес Т.А., Элиас В.П., Скалецкий И.К., Гут Б.Э., Родригес Дж.Ф., Пьяцца Р.М., Феррейра Л.К., Мартинес МБ. Диарейная кишечная палочка. Braz J Microbiol. 2016; 47 (Приложение 1): 3–30.
Артикул PubMed PubMed Central Google ученый
Теннант С.М., Таушек М., Аззопарди К., Бигхэм А., Беннетт-Вуд В., Хартланд Е.Л., Ци В., Уиттам Т.С., Робинс-Браун Р.М. Характеристика атипичных энтеропатогенных штаммов E. Coli клинического происхождения.BMC Microbiol. 2009; 9: 117.
Coli клинического происхождения.BMC Microbiol. 2009; 9: 117.
Артикул PubMed PubMed Central Google ученый
Wang X, Wang J, Sun H, Xia S, Duan R, Liang J, Xiao Y, Qiu H, Shan G, Jing H. Этиология детской инфекционной диареи в развитом регионе Китая: по сравнению с диарея у детей в развивающихся регионах и диарея у взрослых в развитых регионах. PLoS One. 2015; 10 (11): e0142136.
Артикул PubMed PubMed Central Google ученый
Цюй М, Дэн И, Чжан Х, Лю Г, Хуан И, Линь Ц, Ли Дж, Ян Х, Ли Х, Цзя Л. и др. Этиология острой диареи, вызванной энтеропатогенными бактериями, в Пекине, Китай. J Infect. 2012; 65 (3): 214–22.
Артикул PubMed Google ученый
 org/ScholarlyArticle»> 17.
org/ScholarlyArticle»> 17.Алихани М.Ю., Хашеми С.Х., Аслани М.М., Фараджния С. Распространенность и паттерны устойчивости к антибиотикам диарейной кишечной палочки, выделенной у подростков и взрослых в Хамедане, западный Иран.Иран J Microbiol. 2013; 5 (1): 42–7.
PubMed PubMed Central Google ученый
Бролунд А., Эдквист П.Дж., Макитало Б., Олссон-Лильеквист Б., Содерблом Т., Визелл К.Т., Гиске К.Г. Эпидемиология Escherichia Coli, продуцирующая бета-лактамазы расширенного спектра действия, в Швеции, 2007-2011 гг. Clin Microbiol Infect. 2014; 20 (6): O344–52.
CAS Статья PubMed Google ученый
Tian L, Zhu X, Chen Z, Liu W., Li S, Yu W., Zhang W., Xiang X, Sun Z. Характеристики бактериальных патогенов, связанных с острой диареей у детей в возрасте до 5 лет: кросс- секционное исследование. BMC Infect Dis. 2016; 16: 253.
Характеристики бактериальных патогенов, связанных с острой диареей у детей в возрасте до 5 лет: кросс- секционное исследование. BMC Infect Dis. 2016; 16: 253.
Артикул PubMed PubMed Central Google ученый
Институт клинических и лабораторных стандартов. Стандарты эффективности тестирования на чувствительность к противомикробным препаратам, двадцать пятое информационное приложение, M100-S25.Уэйн, Пенсильвания: Институт клинической лаборатории; 2015.
Google ученый
Као CY, Wu HM, Lin WH, Tseng CC, Yan JJ, Wang MC, Teng CH, Wu JJ. Опосредованные плазмидами детерминанты устойчивости к хинолонам у устойчивой к хинолонам Escherichia Coli, выделенной от пациентов с бактериемией в университетской больнице на Тайване, 2001-2015 гг. Научный доклад 2016; 6: 32281.
Научный доклад 2016; 6: 32281.
CAS Статья PubMed PubMed Central Google ученый
Мойо SJ, Gro N, Matee MI, Kitundu J, Myrmel H, Mylvaganam H, Maselle SY, Langeland N. Возрастные этиологические агенты диареи у госпитализированных детей в возрасте до пяти лет в Дар-эс-Саламе, Танзания. BMC Pediatr. 2011; 11:19.
Артикул PubMed PubMed Central Google ученый
Хуанг Ф., Дэн И, Цюй М., Лю Г.Р., Лю И, Чжан Х, Ли Дж., Янь Штаб, Гао Цзы, Лю Б.В. и др. Этиологический надзор и анализ инфекционной диареи в Пекине в 2010 г.Чжунхуа ю фан и сюэ дза чжи [Китайский журнал профилактической медицины]. 2011; 45 (9): 820–4.
Google ученый
 org/ScholarlyArticle»> 24.
org/ScholarlyArticle»> 24.Neter E, Westphal O, Luderitz O, Gino RM, Gorzynski EA. Демонстрация антител к энтеропатогенной Escherichia Coli в сыворотках крови детей разного возраста. Педиатрия. 1955. 16 (6): 801–8.
CAS PubMed Google ученый
Patzi-Vargas S, Zaidi MB, Perez-Martinez I, Leon-Cen M, Michel-Ayala A, Chaussabel D, Estrada-Garcia T.Диарейные бактерии Escherichia Coli, несущие дополнительные гены вирулентности, являются важной причиной умеренных и тяжелых диарейных заболеваний в Мексике. PLoS Negl Trop Dis. 2015; 9 (3): e0003510.
Артикул PubMed PubMed Central Google ученый
Бохари Х., Шах М.А., Асад С., Ахтар С., Акрам М., Рен Б.В. Патотипы Escherichia Coli в Пакистане в результате последовательных наводнений в 2010 и 2011 годах. Am J Trop Med Hyg. 2013. 88 (3): 519–25.
Am J Trop Med Hyg. 2013. 88 (3): 519–25.
CAS Статья PubMed PubMed Central Google ученый
Каур П., Чакраборти А., Асеа А. Энтероагрегантная Escherichia Coli: новый кишечный возбудитель пищевого происхождения. Междисциплинарная перспектива Infect Dis. 2010; 2010: 254159.
CAS Статья PubMed PubMed Central Google ученый
Foster MA, Iqbal J, Zhang C, McHenry R, Cleveland BE, Romero-Herazo Y, Fonnesbeck C, Payne DC, Chappell JD, Halasa N, et al.Энтеропатогенная и энтероагрегантная E. Coli в стуле у детей с острым гастроэнтеритом в округе Дэвидсон, штат Теннесси. Diagn Microbiol Infect Dis. 2015; 83 (3): 319–24.
Артикул PubMed PubMed Central Google ученый
 org/ScholarlyArticle»> 29.
org/ScholarlyArticle»> 29.Кармали MA. Возникающие проблемы общественного здравоохранения, связанные с продуцирующей токсин шига Escherichia Coli, связаны с изменениями в патогене, населении и окружающей среде. Clin Infect Dis. 2017; 64 (3): 371–6.
Артикул PubMed Google ученый
Нахджавани Ф.А., Эманейни М., Хоссейни Х., Иман-Эйни Х., Алиголи М., Джабаламели Ф., Хаги-Аштиани М.Т., Тахерикалани М., Мирсалехиан А. Молекулярный анализ типичных и атипичных энтеропатогенных бактерий Escherichia Coli. от детей с диареей. J Med Microbiol. 2013; 62 (Pt 2): 191–5.
CAS Статья PubMed Google ученый
Чжу XH, Tian L, Cheng ZJ, Liu WY, Li S, Yu W.T., Zhang WQ, Xiang X, Sun ZY. Вирусная и бактериальная этиология острой диареи у детей до 5 лет в Ухане, Китай. Чин Мед Дж. 2016; 129 (16): 1939–44.
Вирусная и бактериальная этиология острой диареи у детей до 5 лет в Ухане, Китай. Чин Мед Дж. 2016; 129 (16): 1939–44.
Артикул PubMed PubMed Central Google ученый
Chen Y, Chen X, Zheng S, Yu F, Kong H, Yang Q, Cui D, Chen N, Lou B, Li X и др. Серотипы, генотипы и паттерны устойчивости к противомикробным препаратам диарейных изолятов кишечной палочки человека, циркулирующих в юго-восточном Китае.Clin Microbiol Infect. 2014; 20 (1): 52–8.
CAS Статья PubMed Google ученый
Lv L, Куропатка SR, He L, Zeng Z, He D, Ye J, Liu JH. Генетическая характеристика плазмид IncI2, несущих blaCTX-M-55, распространяющихся как у домашних, так и у пищевых животных в Китае. Антимикробные агенты Chemother. 2013. 57 (6): 2824–7.
Антимикробные агенты Chemother. 2013. 57 (6): 2824–7.
CAS Статья PubMed PubMed Central Google ученый
Kiratisin P, Apisarnthanarak A, Saifon P, Laesripa C, Kitphati R, Mundy LM. Появление новой устойчивой к цефтазидиму CTX-M бета-лактамазы расширенного спектра, CTX-M-55, как при внебольничных, так и при внутрибольничных инфекциях в Таиланде. Diagn Microbiol Infect Dis. 2007. 58 (3): 349–55.
CAS Статья PubMed Google ученый
Zhang J, Zheng B, Zhao L, Wei Z, Ji J, Li L, Xiao Y. Высокая распространенность CTX-M по всей стране и увеличение CTX-M-55 в Escherichia Coli, выделенных от пациентов с инфекции, возникшие в окружении местных жителей, в районных больницах Китая.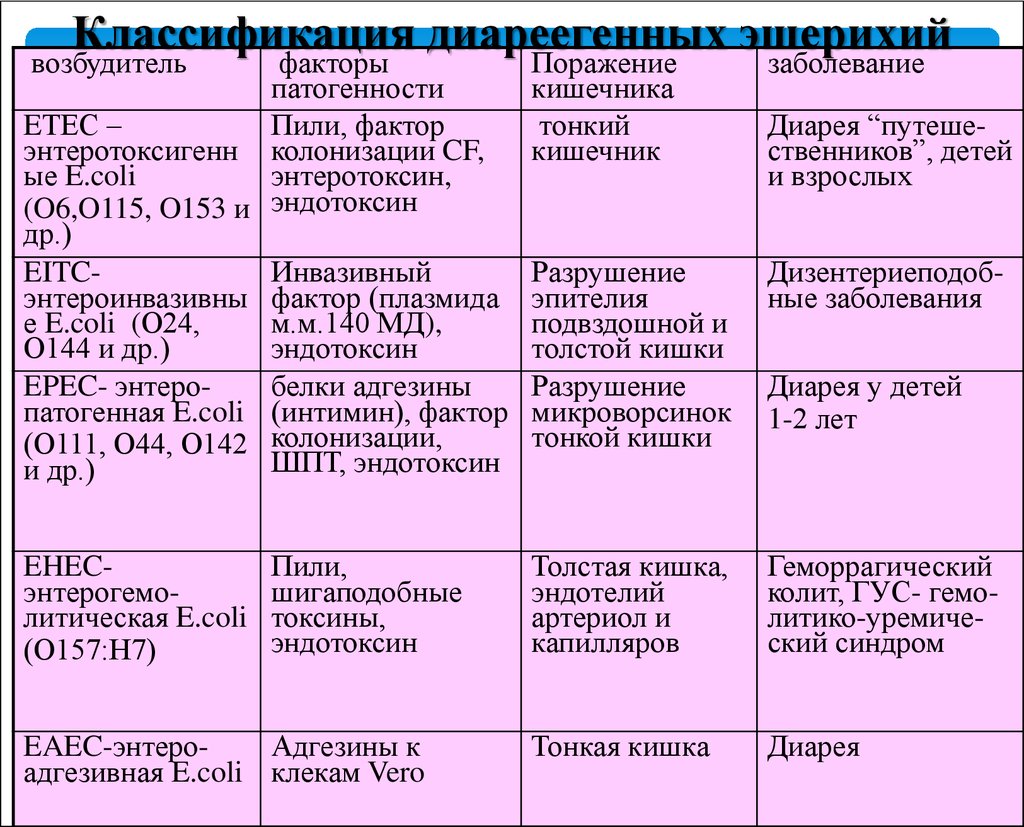 BMC Infect Dis. 2014; 14: 659.
BMC Infect Dis. 2014; 14: 659.
Артикул PubMed PubMed Central Google ученый
Цай JC, Zhang R, Hu YY, Zhou HW, Chen GX. Появление изолятов типа 131 с последовательностью Escherichia Coli, продуцирующих карбапенемазу KPC-2, в Китае. Антимикробные агенты Chemother. 2014. 58 (2): 1146–52.
Артикул PubMed PubMed Central Google ученый
Zhang F, Zhu D, Xie L, Guo X, Ni Y, Sun J.Молекулярная эпидемиология Escherichia Coli, продуцирующая карбапенемазу, и распространенность субклона ST131 h40 в Шанхае, Китай. Eur J Clin Microbiol Infect Dis. 2015; 34 (6): 1263–9.
CAS Статья PubMed Google ученый
 org/ScholarlyArticle»> 38.
org/ScholarlyArticle»> 38.Lou Z, Qi Y, Qian X, Yang W, Wei Z. Появление последовательности типа 131 Escherichia Coli, продуцирующей карбапенемазу Klebsiella Pneumoniae, в Ханчжоу, Китай. Chin Med J. 2014; 127 (3): 528–31.
CAS PubMed Google ученый
Johnson AP, Woodford N. Глобальное распространение устойчивости к антибиотикам: пример устойчивости к карбапенему, опосредованной металло-бета-лактамазой (NDM) в Нью-Дели. J Med Microbiol. 2013; 62 (Pt 4): 499–513.
CAS Статья PubMed Google ученый
Yu J, Tan K, Rong Z, Wang Y, Chen Z, Zhu X, Wu L, Tan L, Xiong W., Sun Z, et al. Нозокомиальная вспышка Klebsiella Pneumoniae, продуцирующих KPC-2 и NDM-1, в неонатальном отделении: ретроспективное исследование. BMC Infect Dis. 2016; 16 (1): 563.
BMC Infect Dis. 2016; 16 (1): 563.
Артикул PubMed PubMed Central Google ученый
Тобиас Дж., Кассем Э., Рубинштейн Ю., Бялик А., Вутукуру С.Р., Наваро А., Рокни А., Валински Л., Эфрос М., Коэн Д. и др. Вовлечение основной диареи, вызывающей кишечную палочку, с упором на энтероагрегантную кишечную палочку, в тяжелую неэпидемическую педиатрическую диарею в стране с высоким уровнем доходов. BMC Infect Dis. 2015; 15:79.
Артикул PubMed PubMed Central Google ученый
Ху Дж., Торрес АГ. Энтеропатогенная кишечная палочка: враг или невиновный наблюдатель? Clin Microbiol Infect. 2015; 21 (8): 729–34.
CAS Статья PubMed PubMed Central Google ученый
 org/ScholarlyArticle»> 43.
org/ScholarlyArticle»> 43.Очоа Т.Дж., Барлетта Ф., Контрерас С., Меркадо Е. Новые сведения об эпидемиологии энтеропатогенной инфекции, вызванной Escherichia Coli. Trans R Soc Trop Med Hyg. 2008. 102 (9): 852–6.
Артикул PubMed PubMed Central Google ученый
Hernandes RT, Elias WP, Vieira MA, Gomes TA. Обзор атипичной энтеропатогенной Escherichia Coli. FEMS Microbiol Lett. 2009. 297 (2): 137–49.
CAS Статья PubMed Google ученый
Хао Р, Цю С., Ван И, Ян Г, Су В, Сун Л., Чжан Дж, Чен Дж, Цзя Л., Ван Л. и др. Устойчивый к хинолонам Escherichia Coli O127a: серотип K63 с фенотипом бета-лактамазы расширенного спектра, вызванный вспышкой пищевого отравления в Китае.
